Team
Team leader: Kiss Tamas
Presentation
The nuclei of human cells contain hundreds of small nuclear regulatory RNAs (snRNAs) which, in forms of small nuclear ribonucleoproteins (snRNPs), function in all aspects of gene expression. In the past, our group has made several fundamental discoveries which largely contributed to our current understanding of the structural and functional complexity of human snRNPs. Currently, two independent research projects are running in our laboratory. One of these programs focuses on dissection of the functional diversity and molecular dynamics of human box C/D and H/ACA modification guide snRNAs which direct site specific 2’-O-ribose methylation and pseudouridylation of ribosomal and spliceosomal RNAs. The other project is aimed at dissection of the complex transcriptional regulatory function of the 7SK snRNP that controls the elongation capacity of RNA polymerase II.
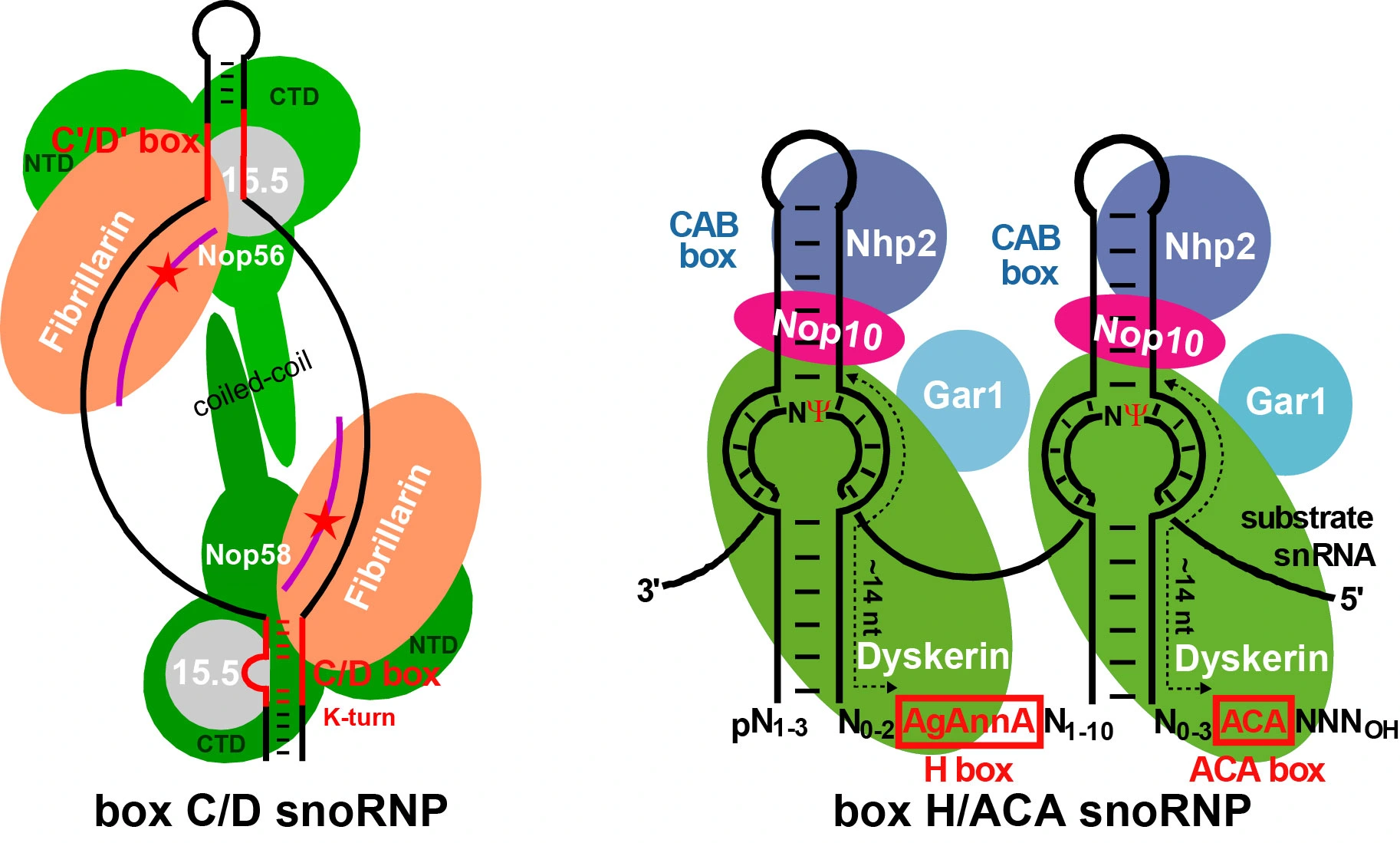
Project 1
Site-specific 2’-O-ribose methylation and pseudouridylation of ribosomal RNAs (rRNAs) and spliceosomal snRNAs is directed by box C/D 2’-O-methylation and box H/ACA pseudouridylation guide RNPs. Each guide RNP is composed of a specific guide RNA and a set of common C/D or H/ACA RNP proteins, including the fibrillarin 2’-O-methyltransferase and the dyskerin pseudouridine synthase. The guide RNPs function through formation of transient base-pairing interactions with their target RNAs. This interaction positions the substrate nucleotide at the catalytic center of the guide RNA-associated modification enzyme. However, some box C/D and H/ACA RNAs lack base-pairing capacity to select rRNA or snRNA sequences for 2’-O-methylation and pseudouridylation. The function of these, so called “orphan” guide RNAs remains unknown. Currently, we are using in vivo RNA-RNA crosslinking-based techniques to identify target RNAs for “orphan” guide RNAs. Earlier, we have demonstrated that some box H/ACA guide RNAs are able to form dynamic alternative interactions with their target sequences in order to position two closely located substrate uridines for pseudouridylation. Recently, we have demonstrated that some H/ACA guide RNS can form alternative interactions with their target RNAs to direct pseudouridylation of two substrate uridines. Now, we investigate whether “guide RNA acrobatics’’ could also contribute to box C/D RNA-directed 2’-O-methylation of human rRNAs and snRNAs.
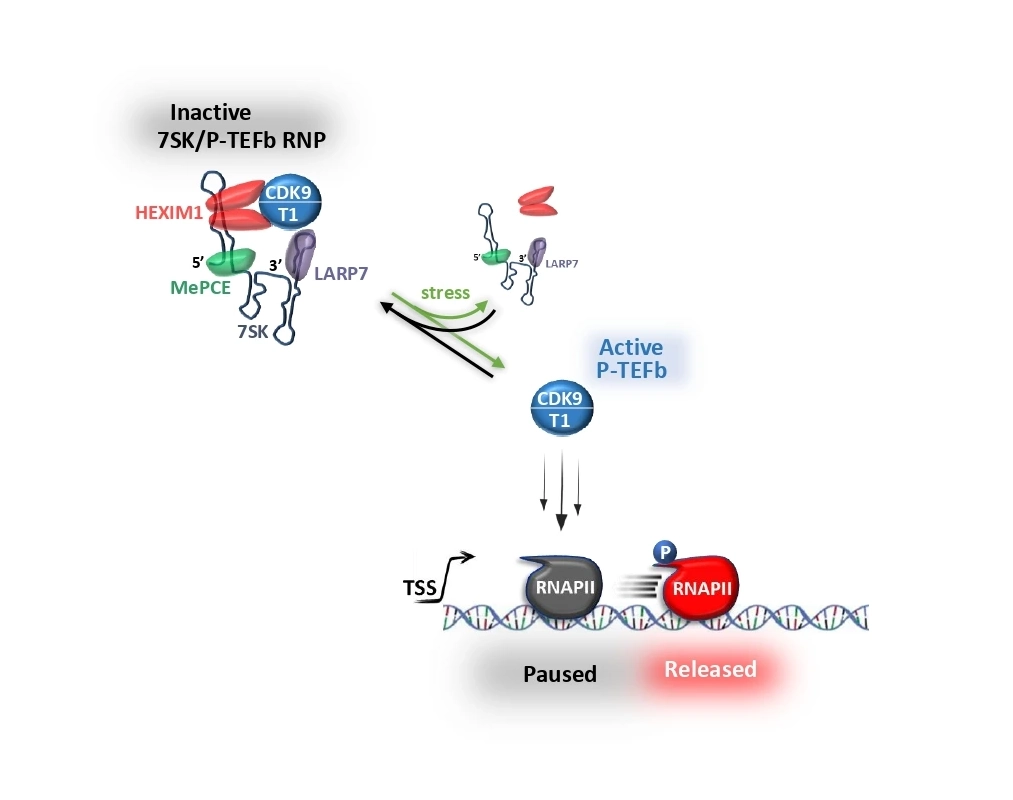
Project 2
Promoter-proximal pausing is a key step in the regulation of protein-coding gene transcription by RNA polymerase II (RNAPII). The transition to productive elongation relies on positive transcription elongation factor b (P-TEFb), a kinase that promotes the synthesis of full-length messenger RNAs by releasing paused RNAPII into the gene bodies. P-TEFb activity is primarily controlled by the 7SK snRNA which, together with the MePCE, LARP7 and HEXIM1 proteins, sequesters P-TEFb into a large kinase-inactive 7SK/P-TEFb snRNP. The 7SK snRNP controls the availability of P-TEFb by releasing or capturing active P-TEFb according to the transcriptional needs of the cell, making it potentially one of the most effective ways of controlling RNAPII pausing in human cells. For example, P-TEFb is rapidly dissociated from the 7SK snRNP during stress response, leading to the genome-wide release of paused RNAPII and enabling adaptive reprogramming of transcription. Thus, the 7SK snRNP may control the timing of key transcriptional transitions, not only during stress response, but also during development and tumorigenesis. Indeed, impaired binding of 7SK snRNP to P-TEFb leads to malignant transformation of cultured cells and has been associated with various cancers, cardiac hypertrophy, neurodevelopmental disorders, as well as inflammatory and autoimmune diseases. Understanding of the complex transcriptional regulatory function of human 7SK snRNP in various physiological and pathological contexts is the major objective of our work.