
Team
Team leader: Trouche Didier
Presentation
Chromatin modifying enzymes and chromatin modifications integrate signals produced by extra or intra-cellular environmental changes in order to adapt gene expression and establish the correct cellular response. In addition, they participate in the maintenance of genetic information during mammalian cells proliferation. As such, chromatin modifying enzymes and chromatin modifications play major role in diseases such as cancers, which are characterized by an increased proliferation rate and by a high genetic instability.
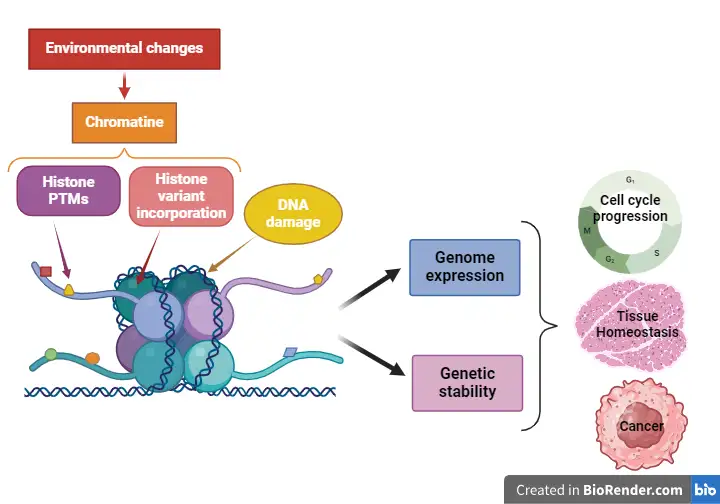
Our team is interested in the function of chromatin modifications in the regulation of genome expression and genetic stability, thereby bringing about the correct control of processes such as cell cycle progression and cell fate. More specifically, we are studying the function of the histone variant H2A.Z in transcription, co-transcriptional mRNA processing and genetic stability during cell cycle progression. We are also investigating the management of oxidative DNA damage in the context of chromatin and how it affects chromatin structure and function. Finally, we are also studying the relationship between chromatin structure and the speed of RNA pol II, focusing on mechanisms linked to transcription elongation, termination and co-transcriptional processing of mRNA.
Project 1
TROUCHE Didier, DR1 CNRS
CANITROT Yvan, DR2 CNRS
BERGOGLIO Valérie, CRCN CNRS
CHAILLEUX Catherine, IRCN CNRS
CINTORI Luana, Doctorante
The general theme of our work is the study of factors influencing the various DNA repair mechanisms and genetic stability in response to DNA damage. Our approach focuses on the role of chromatin and factors that affect its dynamics. Through the use and development of original cellular tools, and using cellular, molecular and imaging approaches as well as genomics data, we are attempting to dissect the choreography involved in DNA damage management in order to better understand the consequences of their defects, with the aim of providing new therapeutic targets for improving the efficacy of treatments that target DNA in cancers.
We are currently focusing on oxidative DNA damage, which are the most common form of DNA damage. How these damages are managed in various chromatin landscapes and how they affect chromatin function is still poorly understood given that most studies used oxidative treatments which induce DNA damage randomly throughout the genome. In addition, these treatments lead to many other oxidative damages than damages to DNA.
We developed opto-genetic tools which allow the targeted generation of oxidative DNA damage. With this original system in hands, we are studying whether specific chromatin landscapes modulate differentially the management of oxidative DNA damage, and thus whether some genomic loci are more prone to oxidative DNA damage-induced mutagenesis. We are also investigating the consequences of oxidative DNA damage on chromatin function, ie transcription and transcription-linked processes.
Selected publications: Fages et al., 2020 ; Taty-Taty et al., 2016 ; Courilleau et al., 2012, Matterra et al., 2010.
Project 2
TROUCHE Didier, DR1 CNRS
MUNIZ Lisa, CRCN CNRS
IACOVONE Marika, Doctorante
EISELE Maurane, Doctorante
While genetic information is the same in every cell of the organism, the expression of this information is specific to the cell’s function and role. The first step in the expression of genetic information, called transcription and carried out by RNA polymerases, consists in reading the information contained in a gene to produce an RNA (Ribonucleic Acid). Mammalian cells have three different RNA polymerases (called I, II and III) which are specialized in the synthesis of the various RNAs essential to the life of the cell. Messenger RNAs, which code for proteins, are synthesized by RNA polymerase II.
Transcription by RNA polymerase II has been extensively studied, but one of the parameters of this process, the speed of elongating RNA pol II (i.e. the speed at which the RNA polymerase moves over the DNA to read the information contained in a gene, thus regulating the number of nucleotides added to the RNA per unit of time), remains poorly characterized. It is known, however, that this speed of transcription is highly dynamic and extremely regulated both within individual genes and between genes. Moreover, the speed of transcription by RNA pol II is a major determinant of the identity of RNAs produced from a gene. Indeed, studies using RNA pol II mutants transcribing faster or slower have shown that the speed of transcription by RNA pol II can control splicing, production of circular RNAs, alternative polyadenylation and transcription termination. We have recently shown that a localized change in RNA pol II transcription speed occurs downstream of some genes in response to oncogenic stress, and that this correlates with an increase in read-through transcription downstream of the gene. Our work thus suggests that localized changes in RNA pol II velocity upon commitment in a particular cell fate could cause changes in transcriptome. However, nothing is known about the factors that regulate locally the speed of RNA pol II.
We have constructed a reporter system allowing to measure changes in RNA polymerase II speed. We used this system to perform a CRISPR/Cas9-based screening to identify factors potentially regulating RNA pol II speed. We are now characterizing the function of chromatin modifying enzymes that came out from this screen in regulating RNA pol II speed at specific loci in the genome.
Selected publications : Muniz et al., 2021a, Muniz et al., 2021b, Muniz et al., 2017
Project 3
TROUCHE Didier, DR1 CNRS
FERNANDEZ-VIDAL Anne, MCUCN CNRS
ESCAFFIT Fabrice, CRHC CNRS
REYES Céline, IECN UT3-PS
IACOVONE Marika, Etudiante en thèse
The nucleosome, composed of an octamer of 4 histones around which DNA is wound, is the basic unit of chromatin. It can undergo major qualitative modifications such as post-translational histone modifications and/or the incorporation of histone variants in place of canonical histones at precise positions in the genome, thus regulating major chromatin functions. The histone variant H2A.Z is one of three histone variants conserved from yeast to human. In vertebrates, H2A.Z is encoded by two genes, giving rise to two isoforms, H2A.Z.1 and H2A.Z.2, and a splice variant, H2A.Z.2.2.
H2A.Z is enriched around the transcription start site of active genes and it plays major roles in the control of transcription of specific genes, therefore regulating cell fate. H2A.Z also appears to be required for the maintenance of genomic information. Indeed, in addition to a still controversial role in DNA double-strand break repair, a function for H2A.Z in Mitosis and S-phase DNA replication has been suggested. Importantly, H2A.Z isoforms or their regulators have been found overexpressed or mutated in numerous cancers.
Our aim is to characterize the function of H2A.Z isoforms in genome expression, cell fate control and genetic stability.
To overcome the limitations underlying the use of siRNAs, we have constructed by genome editing, cell lines enabling us to induce the degradation of endogenous H2A.Z isoforms in a rapid, inducible and reversible manner. We used these cell lines to perform CRIPR/Cas9 genome wide screening to identify genes synthetic lethal with H2A.Z isoforms, leading to the identification of many pertinent pathways. We have also generated cell lines to track the localization of endogenous H2A.Z in real time and cell lines with different abilities to commit in different fates. We are now using these cell lines to characterize the direct function of H2A.Z isoforms in transcription, cell cycle progression, DNA repair and cell fate control.
Selected publications : Rispal et al., 2024, Lamaa et al., 2020, Rispal et al., 2019, Lazorthes et al., 2015
Team members
– Rispal J, Rives C, Jouffret V, Leoni C, Dubois L, Chevillard-Briet M, Trouche D, Escaffit F. Control of intestinal stemness and cell lineage by histone variant H2A.Z isoforms (2024) Mol Cell Biol. 18:1-18.
– Ouvrard J, Muniz L, Nicolas E, Trouche D. (2022) Small Interfering RNAs Targeting a Chromatin-Associated RNA Induce Its Transcriptional Silencing in Human Cells. Mol Cell Biol. 42(12):e0027122
– Muniz L, Nicolas E, Trouche D. (2021) RNA pol II speed: a key player in control and adaptation of transcriptome composition, EMBO J. 40(15): e105740.
– Muniz L, Lazorthes S, Delmas M, Ouvrard J, Aguirrebengoa M, *Trouche D, *Nicolas E (* co-last authors) (2021). Circular AnRIL RNAs switch from repressor to activator of p15/CDKN2B expression during RAF1 oncogene-induced senescence. RNA Biol. 10.1080.
– Fages J, Chailleux C, Humbert J, Jang S-M, Loehr J, Lambert J-P, Coté J, *Trouche D, *Canitrot Y (* co-last authors). (2020) JMJD6 participates in the maintenance of ribosomal DNA integrity in response to DNA damage. PLoS Genetics, 16(6): e1008511.
– Lamaa A, Humbert J, Aguirrebengoa M, Xue C, Nicolas E, Coté J, Trouche D. (2020) Integrated analysis of H2A.Z isoforms functions reveals a complex interplay in gene regulation. eLife 28 (9), doi: 10.7554/eLife.53375.
– Rispal J, Baron L, Beaulieu J-F, Chevillard-Briet M, *Trouche D, *Escaffit F (*co-last authors) (2019) The H2A.Z histone variant integrates Wnt signaling in intestinal epithelial homeostasis. Nature Comm 10 (1): 1827.
– §Muniz L., §Deb M.K. (§co-first authors), Aguirrebengoa M, , Lazorthes S., *Trouche D, *Nicolas E. (* co-last authors) (2017) Control of gene expression in senescence through transcriptional read-through of convergent protein-coding genes. Cell Reports 21(9):2433-2446.
– Grezy, A., Chevillard-Briet, M., *Trouche, D., *Escaffit, F. (* co-last authors) (2016) Control of genetic stability by a new heterochromatin compaction pathway involving the Tip60 histone acetyltransferase. Mol Biol Cell 15: 599-607.
– Taty-Taty, G., Chailleux, C., Quaranta, M., So, A., Guirouilh-Barbat, J., Lopez, B., Bertrand, P., Trouche, D., Canitrot, Y. (2016) Control of alternative end joining by the chromatin remodeler p400 ATPase. Nucleic Acids Res 44: 1657-1668.
– Salifou, K, Ray, S, Verrier, L., Aguirrebengoa, M., Trouche, D., *Panov, K.I., *Vandromme, M. (* co-last authors) (2016) The histone demethylase JMJD2A/KDM4A links nutrients/growth factors availability and ribosomal RNA transcription. Nature Comm. 7: 10174.
– Lazorthes, S., Vallot, C. Briois, S., Aguirrebengoa, M, Thuret, J.-Y., St. Laurent, G., Rougeulle, C., Kapranov, P., Mann, P., *Trouche, D, *Nicolas, E (* co-last authors). (2015) A vlincRNA participates in senescence maintenance by relieving H2AZ-mediated repression at the INK4 locus in trans, Nature Comm. 6:5971.
– Chevillard-Briet, M., Quaranta, M., Grezy, A., Matterra, L., Courilleau, C., Philippe, M., Mercier, P., Corpet, D., Lough, J., Ueda, T., Fukunaga, R., *Trouche, D., *Escaffit, F. (*co-last authors). (2014) Interplay between chromatin modifying enzymes controls colon cancer progression through Wnt signaling. Human Mol Genet 23: 2120-31.
– Courilleau, C., Chailleux, C., Jauneau, A., Grimmal, F., Briois, S., Boutet-Robinet, E., Boudsocq, F., *Trouche, D., *Canitrot, Y. (*co last-authors) (2012) The chromatin remodeler p400 ATPase facilitates Rad51 mediated repair of DNA double-strand breaks. J Cell Biol, 199: 1067-1081.
– Verrier, L., Escaffit, F., Chailleux, C., *Trouche, D., *Vandromme, M. (* co-last-authors). (2011) A new isoform of histone demethylase JMJD2A/KDM4A required for skeletal muscle differentiation. Plos Genet. 2011 7(6):e1001390.
– Capp, J.P., Boudsocq, F., Bergoglio, V., Trouche, D., Cazaux, C., Blanco, L., Hoffmann, J.S., Canitrot, Y. (2010) The R438W polymorphism of human DNA polymerase lambda triggers sensitivity to camptothecin by compromising the homologous recombination pathway. Carcinogenesis, 31(10): 1742-7.
– *Mattera, L., *Courilleau, C. (*co-first authors), Legube, G., Ueda, T., Fukunaga, R., Chevillard-Briet, M., §Canitrot, Y., §Escaffit, F. (§ equal contribution), Trouche, D. (2010) The E1A-associated p400 protein modulates cell fate decision by the regulation of ROS homeostasis. PLos Genet. 6(6):e1000983.
– *Chailleux, C., *Tyteca, S. (*co-first authors), Courilleau, C., Papin, C., Boudsocq, F., Puget, N., Grigoriev, M., Canitrot, Y., Trouche, D. (2010) Physical interaction between the histone acetyl transferase Tip60 and the DNA double strand breaks sensor MRN complex. Biochem J., 426: 365-371.
– *Mattera, L., *Escaffit, F. (*co-first authors), Pillaire, M.-J., Selves, J., Tyteca, S., Hoffmann, J.-S., Gourraud, P.-A., §Chevillard-Briet, M., §Cazaux, C. (§ equal contribution), Trouche, D. (2009) The p400/Tip60 ratio is critical for colorectal cancer cells proliferation through DNA damage response pathways. Oncogene, 28: 1506-1517.
– Tyteca, S., Vandromme, M., Legube, G., Chevillard-Briet, M., Trouche, D. (2006) Tip60 and p400 are both required for UV-induced apoptosis but play antagonistic roles in cell cycle progression. EMBO J., 25: 1680-1689. Selected by EMBO J editors as one of the four best papers published by EMBO J during the first half of 2006 (EMBOencounters 6, summer 2006)
– *Daury, L., *Chailleux, C. (*co-first authors), Trouche, D. (2006) Deposition of histone H3.3 on E2F responsive genes is linked to transcription. EMBO R. 7: 66-71. Régnier, V. (corresponding author), Vagnarelli, P., Fukagawa, T., Zerjal, T., Burns, E., Trouche, D., Earnshaw, W., Brown, W. R. (2005) CENP-A is required for accurate chromosome segregation and sustained kinetochore association of BubR1. Mol. Cell. Biol. 25: 3967-3981.
– Legube, G., *Linares, L.K., *Tyteca, S. (*co-second athors), Caron, C., Scheffner, M., Chevillard-Briet, M., and Trouche, D. (2004) Role of the histone acetyl transferase Tip60 in the p53 pathway. J. Biol. Chem. 279: 44825-44833.
– Legube, G., Linares, L.K., Lemercier, C., Scheffner, M., Khochbin, S., and Trouche, D. (2002) The histone acetyl transferase Tip60 is targeted to proteasome-mediated degradation by the Mdm2 oncoprotein. EMBO J. 21:1704-1712.
Affiliation




