
Team
Team leader: Suzanne Magali
Presentation
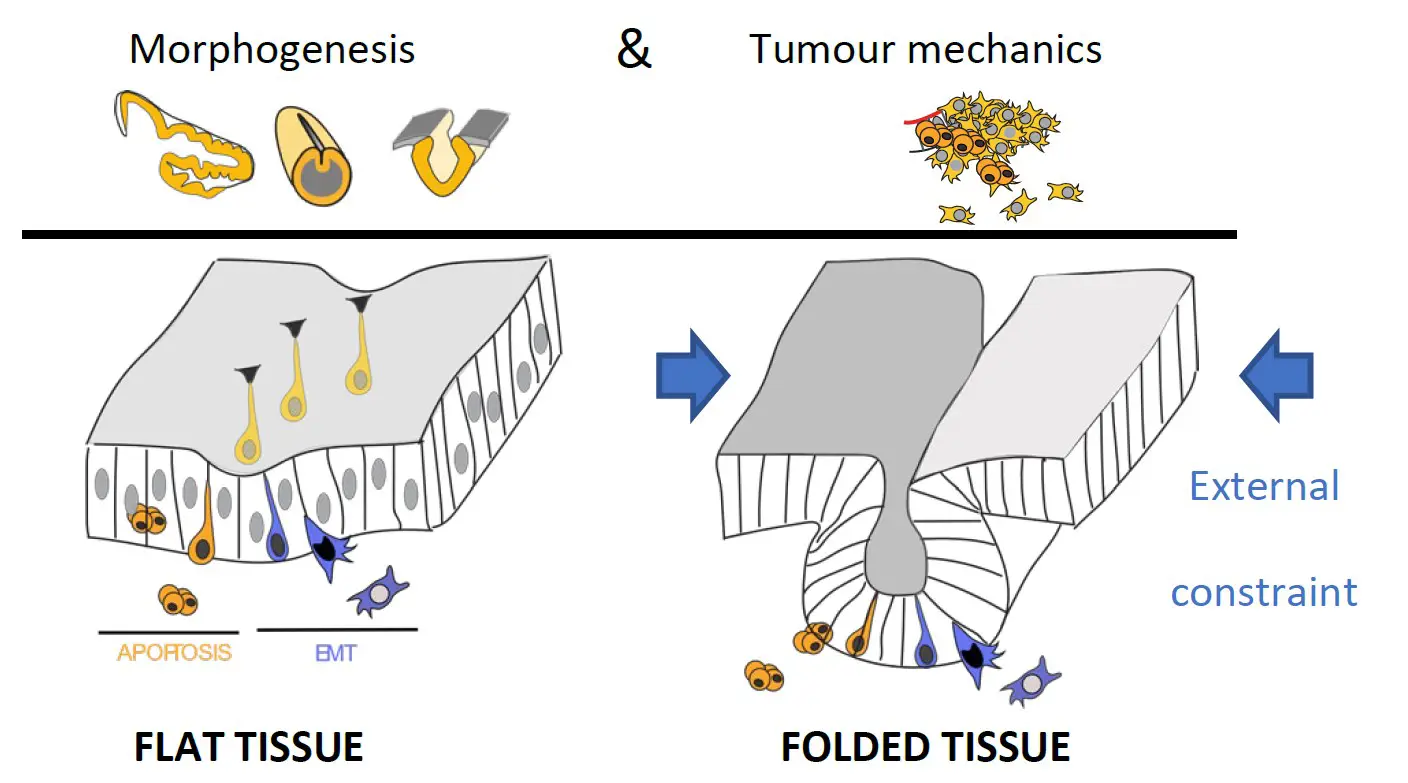
The physics of life is a growing field of research addressing the contribution of physical parameters to living organisms, but also their capability to respond to physical challenges. It is now well recognized that tissue mechanics is not only an integral part of development and tissue homeostasis but is also involved in tumor development. Mechanobiology approaches in vitro have led to fundamental discoveries on the amazing plasticity of cells revealing the importance of mechanical cues in living systems, however, how relevant these observations are in vivo in an integrated model and how cells coordinate their behavior to create new shape is far from being understood.
To better understand how mechanics contribute to tissue plasticity, the team addresses the following questions:
– how cell extrusion, including apoptosis and EMT, are driven mechanically
– how cell extrusion and external constraint influence tissue morphogenesis
– how cell and tissue mechanics influence tumor evolution
These aims are addressed by experimental approaches, using integrated model systems, including drosophila and birds, offering a large set of genetic tools and the possibility to image living tissues as well as by theoretical biophysical modeling to understand the fundamental principles driving cell extrusion and epithelial dynamics.
Project 1
Apoptosis as an active player in morphogenesis
Pioneer studies on apoptosis reveal the surprising importance of cell death during normal development in a number of different organs and organisms (Fuchs Y. and Steller H., 2015). Initially, apoptosis was simply viewed as a way to get rid of cells without considering its potential influence on the surrounding tissue. More recently, it has been suggested that apoptotic cells play an important dynamic role during morphogenesis (Toyama Y. et al., 2008; Suzanne M. et al., 2010; Ambrosini A. et al., 2016). Nonetheless, the mechanisms whereby dying cells drive tissue shape modification was unknown. Using the Drosophila developing leg as a model system to study apoptosis-dependent epithelium folding (Manjon C. et al., 2007), we showed that apoptotic cells, which appear specifically in the fold domain, before and during its formation (see movie 1), produce a pulling force through the unexpected maintenance of their adherens junctions. This force leads to a non-autonomous increase in tissue tension and apical constriction of surrounding cells, leading to epithelium folding. These results reveal that, far from being passively eliminated as generally thought, dying cells are very active until the end of the apoptotic process (Monier B. et al, Nature, 2015).
Movie 1. – Magali Suzanne – Apoptotic dynamics in the developing leg. Apoptotic cells are stained with acridine orange (green), while the apical surface of the epithelium is visualized thanks to DlgGFP (in green too).
Based on this discovery, our objective was to understand how apoptotic cells influence their surroundings from the micro-environment to the macro-scale level and characterize this new regulator of mechanical forces in morphogenesis. To do so, we dissected the cellular mechanisms governing the generation of the apoptotic force and identified network of genes orchestrating apoptosis dependent morphogenesis through a genetic screen.
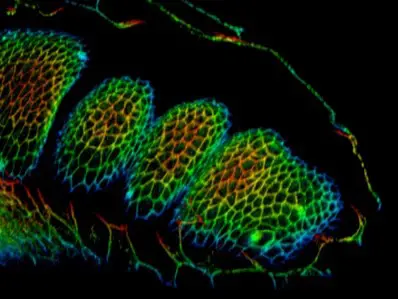
Model used to screen for genes involved in apoptosis-dependent morphogenesis.
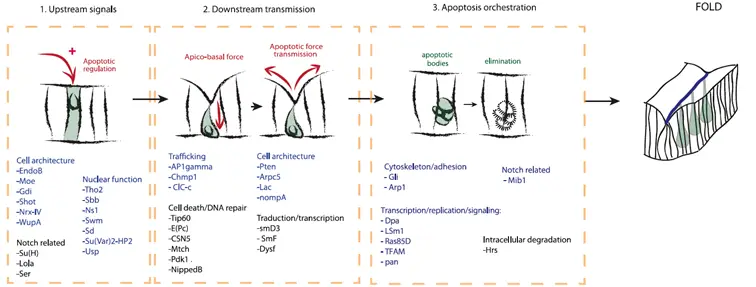
Scheme summarizing the different types of candidates identified through a genetic screen, including: (1) potential new regulators of endogenous apoptosis, (2) potential genes involved in downstream transmission of the apoptotic signal, and (3) genes involved in apoptosis orchestration leading to fold formation.
Evolutive conservation of apoptosis mechanics and impact on morphogenesis in vertebrates
Our work on the drosophila leg indicates that apoptosis can actively contribute to morphogenesis, revealing that apoptotic cells can impact their surroundings. However, how this applies to vertebrate morphogenesis remained unknown. More recently, we used the formation of the neural tube, an initially flat epithelium that bends and fuses (see movie 2), to determine how apoptosis contributes to morphogenesis in vertebrates. Neural tube closure defects have been reported when apoptosis is impaired in chicken and mouse embryos, although the cellular mechanisms involved are unknown. Using chicken and quail embryos, we found that apoptosis mainly occurs shortly before or at the time of dorsal bending of the neural epithelium and is required for this bending. By a combination of high-resolution time-lapse imaging, laser ablation and immunodetection experiments, we show that apoptotic cells generate an apico-basal force before being extruded from the neuro-epithelium. The force generated by apoptotic cells is transmitted to their neighbors, as revealed by concomitant cell deformation observed across several cell diameters, suggesting a local increase in tissue tension. At the subcellular level, this force deforms the nucleus and precipitates its fragmentation. All together these data reveal that apoptotic cells influence their surroundings mechanically and strongly suggest their active contribution to neural tube bending. Furthermore, it suggests that the same force plays a dual role in in tissue morphogenesis and in the orchestration of cellular and nuclear destruction.
Movie 2. – Daniela Roellig – Movie showing the dynamics of neural tube closure in quail embryo
EMT as an active player in morphogenesis
Intriguingly, EMT is often associated with tissue invagination during development; however, the impact of EMT on tissue remodeling remain unexplored. Using the mesoderm invagination (see movie 3) and ectopic induction of EMT in the leg disc in Drosophila, we showed that at the initiation of the EMT process, cells produce an apico-basal force, orthogonal to the surface of the epithelium (see movie 4) that constitutes an important driving force for tissue invagination in Drosophila. When EMT is ectopically induced, cells starting their delamination generate an orthogonal force and induce ectopic folding. Similarly, during mesoderm invagination, cells undergoing EMT generate an apico-basal force through the formation of apico-basal structures of myosin II. Using both laser microdissection and in silico physical modelling, we show that mesoderm invagination does not proceed if apico-basal forces are impaired, indicating that they constitute driving forces in the folding process. Altogether, these data reveal the mechanical impact of EMT on morphogenesis.
Movie 3. – Mélanie Gracia – Movie showing the dynamics of mesoderm invagination in drosophila embryo. Nuclei are color coded with depth.
Movie 4. – Mélanie Gracia – Myosin II dynamics during mesoderm invagination in drosophila embryo. Note the presence of transient apico-basal structures all along the invagination, at the end of apical constriction.
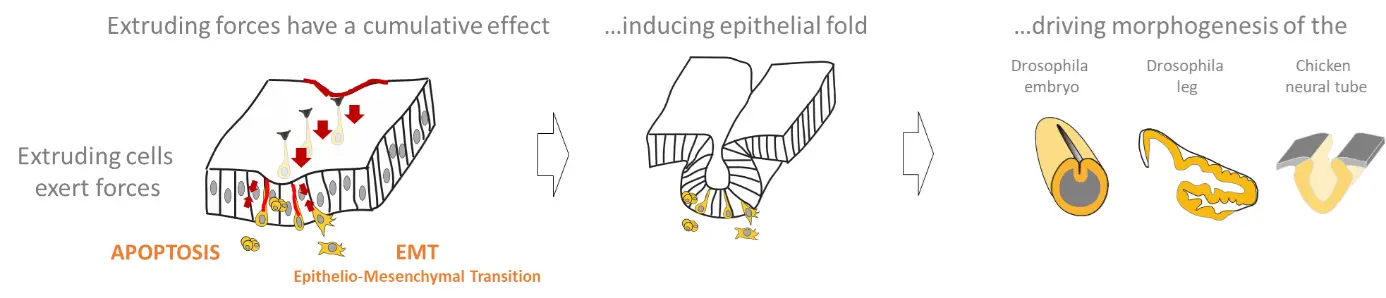
Scheme recapitulating the impact of forces generated by extruding cells on morphogenesis.
Robustness of morphogenesis
We next asked how tissue mechanics controls morphogenetic robustness when challenged by developmental noise. Using Drosophila developing leg and its stereotyped tarsal folds as a model, we show that a bias in force propagation ensures stereotyped morphogenesis despite the presence of mechanical noise in the environment. We found that knockdown of the Arp2/3complex member Arpc5 specifically affects fold directionality while altering neither the developmental nor the force generation patterns (see movie 5). By combining in silico modeling, biophysical tools, and ad hoc genetic tools, our data reveal that junctional myosin II planar polarity favors long-range force channeling and ensures folding robustness, avoiding force scattering and thus isolating the fold domain from surrounding mechanical perturbations.
Movie 5. – Emmanuel Martin – Fold defect in a Arpc5 RNAi context (right), compare to a control leg disc (left). Fold are highlighted in blue and the segmentation pattern of Dpn is in magenta.
Project 2
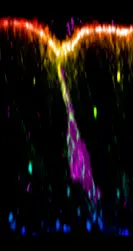
Cellular mechanics of apoptosis
A pioneer study from our lab revealed that apoptotic cells, far from being eliminated passively, actively participate to tissue remodeling. They generate a traction force through the formation of a particular structure of acto-myosin along their apico-basal axis (see movie 1), pull on their neighbors (see movie 2), which increases local tension and drives tissue folding.
Movies 1-2. – Bruno Monier – DE-Cad::GFP (1) and MRLC::GFP (2) dynamics in apoptotic cells (in red in 1) from prefold
stage leg discs. Note the transient formation of an apico-basal structure of myosin II (1) and the deformation of the apical surface (1-2).
We investigated the cellular mechanism responsible for the generation of this apico-basal forces and revealed that the nucleus, on top of its capacity to respond to mechanical forces, constitutes an essential anchoring point for apico-basal force generation, thanks to its stabilization by a basal F-actin network linked to focal adhesions. Then, when the acto-myosin cable contracts, the nucleus basal anchorage is lost and the nucleus goes up (see movie 3). Thus, this work reveals that the nucleus, in addition to its role in genome protection, actively participates in mechanical force production and connects the contractile actomyosin cytoskeleton to basal junctions.
Movie 3. – Megane Rayer – Nucleus (green) and myosin II (red) dynamics during the apoptotic process in the columnar epithelium of the drosophila leg disc.
We further show that this cellular mechanism is conserved in vertebrates using chicken embryos. We show that a tensile myosin II cable-like structure connects the nucleus to the apical surface of the cell and contracts, leading to an upward movement of the nucleus before the fragmentation of the cell (see movie 4).
Movie 4. – Daniela Rolellig – Nucleus (green) and myosin II (magenta) dynamics during the apoptotic process in the columnar epithelium of the chicken neural tube.
Cellular mechanics of EMT
We next wondered how other types of cell extrusion compare to the dynamics observed in apoptotic cells. We turned to epithelial−mesenchymal transition (EMT), an essential process both in physiological and pathological contexts. We found that apico-basal forces are also produced by cells undergoing EMT before their extrusion, similarly to what was observed in apoptotic cells (see figure below), through the formation of an apico-basal structure of myosin II, which is transient. This apico-basal force produces a mechanical signal, which constitutes an important driving force and actively influences the surrounding tissue, as shown by mesoderm invagination failure when this force is prevented.
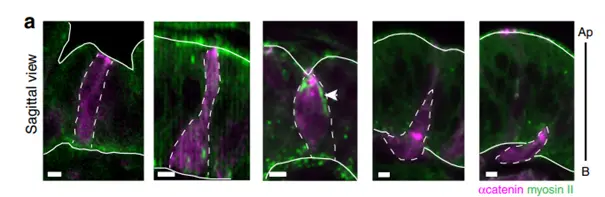
Images of cells undergoing EMT (induced by Snail expression) at different stage of the extrusion process. The EMT cells are in magenta (a-Cat), non-muscular myosin II is in green. Note the presence of an apico-basal transient structure in the central image which coincide with an apical deformation of the epithelium.
Project 3
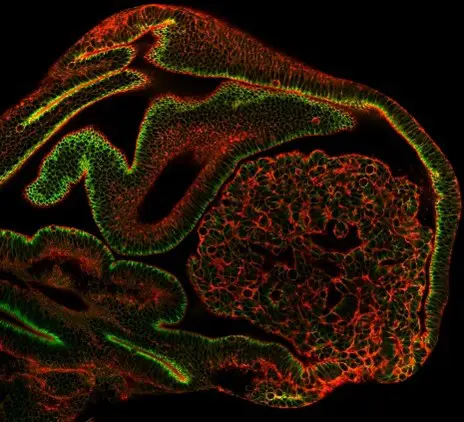
Tumors evolve through the acquisition of increasingly aggressive traits associated with dysplasia. This progression is accompanied by the alteration of tumor mechanical properties, especially through extracellular matrix remodeling. However, the contribution of pretumoral tissue mechanics, especially the subcellular remodeling of cytoskeletal structures, to tumor aggressiveness remains poorly known in vivo.
To decipher how tension evolves during tumor development and how it can impact tumor evolution, we analyzed tissue tension from very early stage of tumor development, even before the apparition of a tumor. We found that adherens junction tension in pre-tumoral tissues dictates subsequent tumor evolution in Drosophila. Increased cell contractility, observed in aggressive tumors before any sign of tissue overgrowth, proved sufficient to trigger dysplasia in normally hyperplastic tumors. Unexpectedly, high contractility precedes any change in cell polarity and contributes to tumor evolution through cell death induction which favors cell-cell junctions weakening. Overall, our results highlight the need to re-evaluate the roles of tumoral cell death, and identify pre-tumor cell mechanics as an unsuspected early marker and key trigger of tumor aggressiveness.
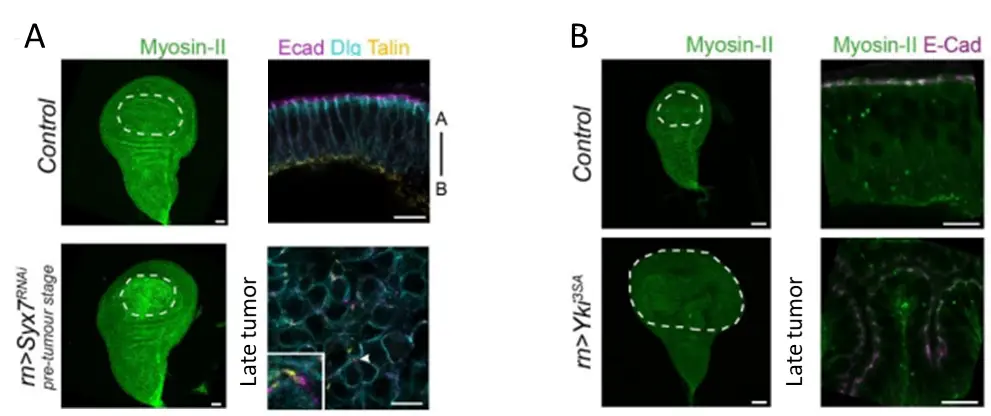
Myosin II accumulates specifically in pretumoral tissues of aggressive tumors. A. Syx7 RNAi pretumoral tissue showing an accumulation of myosin II compared to the control (left), while Syx7 RNAi tissues evolve as dysplastic tumors (right). B. Yki3SA pretumoral tissue shows no sign of myosin II accumulation (left), while Yki3SA tissues evolve as hyperplastic tumors.
Project 4
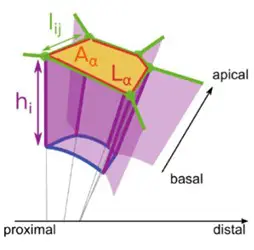
Tyssue: an epithelium simulation library
The Tyssue Python library seeks to provide a unified interface to implement bio-mechanical models of living tissues. Its main focus is on vertex-based epithelium models. Tyssue allows to model the mechanical behavior of 2D, apical 3D or full 3D epithelia based on the numerical resolution of the equations of motion for the mesh vertices. We contribute to the development of this library that has been developed by Guillaume Gay from the Turing Center in Marseille.
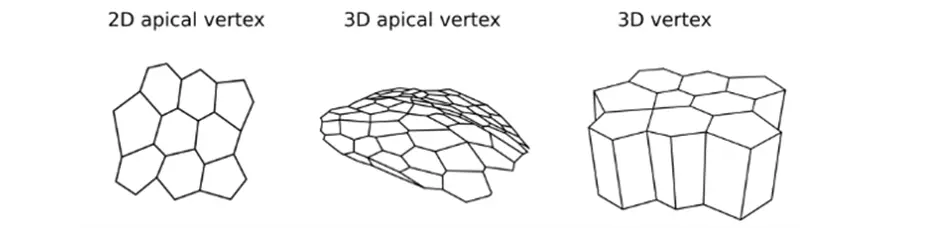
Figure representing the different type of mesh: 2D apical vertex (left), 3D apical vertex (middle) and 3D vertex (right).
Impact of apico-basal apoptotic forces versus apical transmission
To test the role of apoptotic forces in folding, we elaborated a physical model based on the 2D vertex model. In the absence of the apoptotic dependent forces, the model indicates that the simple disappearance of 30 cells from a continuous ring-like domain is not sufficient to induce cell shape reorganization and to create an invagination at the tissue level (see movie 1).However, if a transient apico-basal force is applied in each dying cell, an invagination response is observed all around the cylindrical tissue (see movie 2).This shows that the apoptotic forces are necessary and sufficient to induce folding in silico.
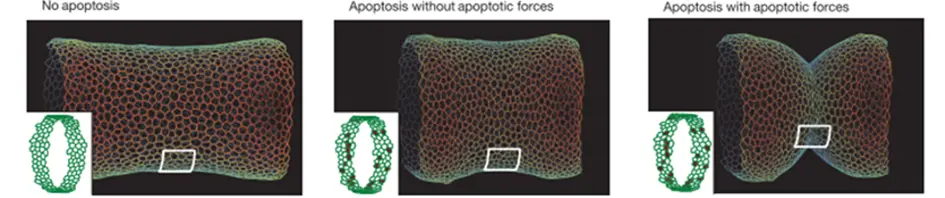
Simulations showing the shape of a cylindrical virtual epithelium with (middle and right) and without (left) the extrusion of around 30 apoptotic cells, and with (right) and without (middle) apoptotic centripetal forces.
Movie 1-2. – Guillaume Gay – Simulations of fold formation with (2) and without (1) centripetal (apico-basal) forces in apoptotic cells.
Morphogenesis Robustness
We observed that developing leg folding occurs in a mechanically noisy environment. To test the impact of mechanical perturbations on fold formation, we implemented our previous model of developing leg folding and integrated random mechanical perturbations in close vicinity to the predicted fold domain, mimicking the mechanical noise observed in vivo. Interestingly, we found that mechanical noise appears sufficient to induce fold deviations in the simulations, mimicking the absence of Myosin planar polarity in the tissue.
We then wondered whether restoring myosin II polarity in a non-polarized tissue could rescue the defects of fold deviation. Although not feasible in vivo, we could address this question using in silico modeling, asking whether fold deviation caused by local mechanical perturbations could be rescued by the introduction of planar polarized junctional myosin II. In order to integrate myosin II planar polarity in the model, we mimicked myosin II planar polarity and the associated tension anisotropy by the attribution of different values of junctional tension depending on junction orientation in our model. Fold robustness was unaffected by the integration of tension polarity in the model in the absence of external perturbations, while interestingly, gradual increase of tissue polarity favors fold straightness and insensitivity to surrounding perturbations. We further quantified fold morphogenetic robustness in our theoretical model for different degrees of polarity. Interestingly, increasing polarity decreases the degree of deviation of the fold and thus decreases the variability in fold directionality, supporting the fact that the planar polarization of tissue tension favors mechanical isolation of fold formation, which ultimately ensures morphogenetic robustness.
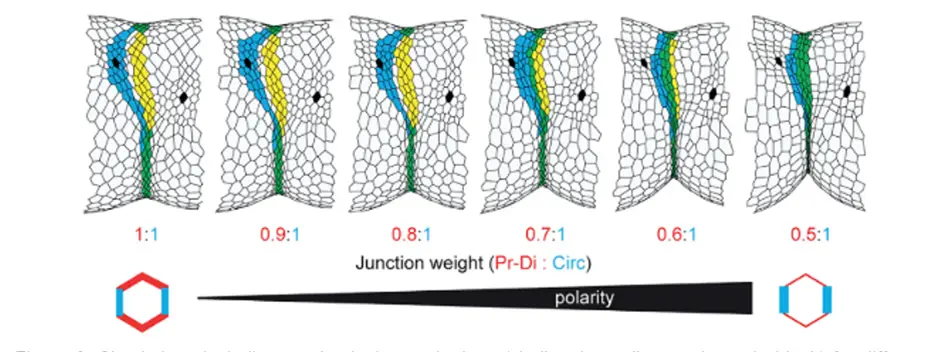
Simulations including mechanical perturbations (challenging cells are shown in black) for different value of junction weight (i.e., tension anisotropy). Predicted fold domain is in yellow, real fold in blue and perfect match in green.
EMT apico-basal forces drive epithelium remodeling
To test specifically the contribution of apico-basal forces generated by EMT cells in tissue folding, we developed a physical model of the embryo apical junction network based on the vertex model we implemented recently. This 3D model of mesoderm invagination mimics the cellular dynamics observed in the embryo (anisotropic constriction, gradual and asynchronous apical constriction, and apical force propagation). In this model, mesodermal cells progressively constrict their apical surface preparing their future delamination. Simulation results showed that apical tension alone without apico-basal force only leads to a curved-shaped mesoderm, independently of the strength of apical contractility. Only in the presence of apico-basal tension does the mesoderm invaginate, forming the V-shape observed in vivo.
Gracia, Theis et al., Nature Communication 2019 doi: 10.1038/s41467-019-10720-0
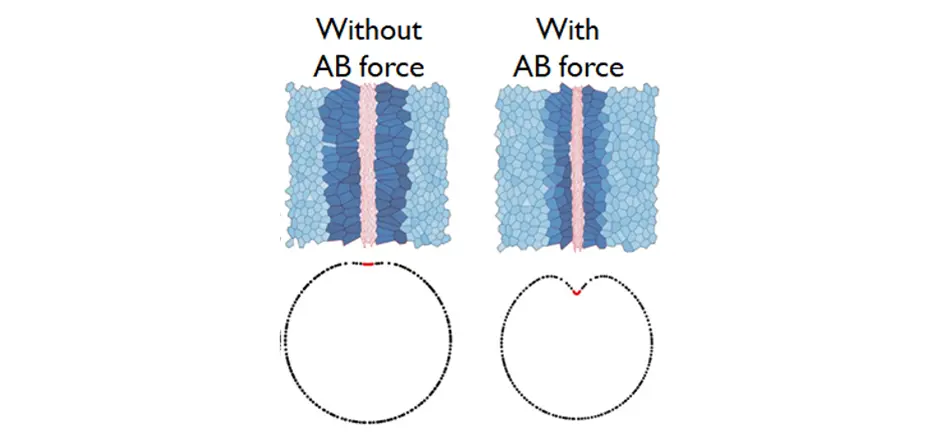
Simulation of mesoderm invagination in absence (left) or presence (right) of apico-basal force.
Project 5
Tools developed in the team
The Apoptosensor: a method to follow apoptotic cell dynamics
In order to characterize apoptotic cell dynamics in the developing leg epithelium, we adapted a method to positively mark apoptotic cells in living tissue to Drosophila. These reporters (based on 3 different fluorophores) allow the specific visualization of apoptotic cells directly within living tissues, without any post-acquisition processing (see movie 1). They also have the advantage to mark apoptotic cells from the initiation of the process to the fragmentation of the cell.
Movie 1. – Magali Suzanne –
DISSECT: A new method to dissect highly deformed epithelial surfaces
Understanding morphogenesis strongly relies on the characterization of tissue topology and mechanical properties deduced from imaging data. An important bottleneck in this field is the need to analyze imaging datasets and extract quantifications not only of cell and tissue morphology but also of the cytoskeletal network’s organization in an automatized way. We developed a method, called DISSECT, for DisPerSE (Discrete Persistent Structure Extractor)-based Segmentation and Exploration of Cells and Tissues, that offers the opportunity to extract automatically, in strongly deformed epithelia, a precise characterization of the spatial organization of a given cytoskeletal network combined with morphological quantifications in highly remodeled three-dimensional (3D) epithelial tissues.
Movie 2. – Sophie Theis –
Movie 3. – Sophie Theis –
Movie 4. – Tatiana Merle –
CONTRIBUTIONS TO TOOL DEVELOPMENT
Tyssue: an epithelium simulation library
The Tyssue Python library seeks to provide a unified interface to implement bio-mechanical models of living tissues. Its main focus is on vertex-based epithelium models. Tyssue allows to model the mechanical behavior of 2D, apical 3D or full 3D epithelia based on the numerical resolution of the equations of motion for the mesh vertices. We contribute to the development of this library that has been developed by Guillaume Gay.
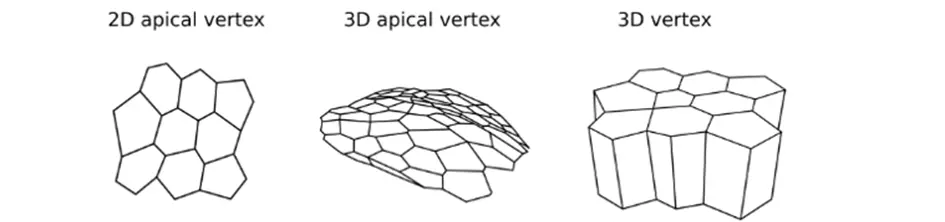
Schemes showing the different geometries of the vertex models: mesh of 2D vertex (left), 3D apical vertex (middle) and 3D vertex (right).
RIM: Super-resolution on living samples
Current super-resolution microscopy (SRM) methods suffer from an intrinsic complexity that might curtail their routine use in cell biology. We contribute to the development of a new method based on random illumination microscopy (RIM), developed by Thomas Mangeat at the CBI, that increases live imaging resolution in x, y and z, avoids tissue optical aberration in depth and is compatible with multicolorlive-cell imaging over extended periods of time.
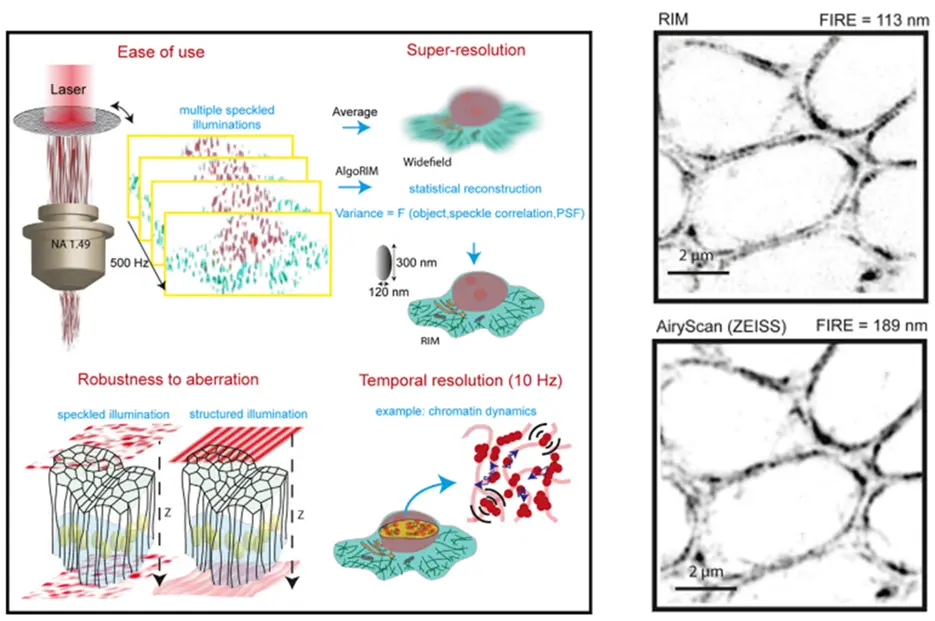
(left) Schematics showing the basic principle of RIM imaging.
(right) Comparison of RIM and Airyscan imaging
Team members
– Merle T.* #, Theis S.*, Kamgoué A., Martin E., Sarron F., Gay G., Farge E. and Suzanne M#. (2023) DISSECT is a tool to segment and explore cell and tissue mechanics in highly deformed 3D epithelia Developmental Cell DOI: 10.1016/j.devcel.2023.07.017
– Schott S., Barbaste A., Benassayag C. and Suzanne M. (2023) Dissecting morphogenetic apoptosis through a genetic screen in Drosophila Life Science Alliance DOI: 10.26508/lsa.202301967
– Roellig D., Theis S., …Suzanne M. (2022) Force-generating apoptotic cells orchestrate avian neural tube bending Developmental Cell doi: 10.1016/j.devcel.2022.02.020
– Martin E., Suzanne M. (2021) mBeRFP: a versatile fluorescent tool to enhance multichannel live imaging and its applications. Development doi: 10.1242/dev.200495.
– Martin E., Theis S., …, Suzanne M. (2021) Arp2/3-dependent mechanical control of morphogenetic robustness in an inherently challenging environment Developmental Cell DOI: https://doi.org/10.1016/j.devcel.2021.01.005
– Gracia M, Theis S, …, Suzanne M (2019) Mechanical impact of epithelial-mesenchymal transition on epithelial morphogenesis in Drosophila. Nat Communication. doi: 10.1038/s41467-019-10720-0
– Ambrosini A, Rayer M, Monier B, Suzanne M. (2019) Mechanical Function of the Nucleus in Force Generation during Epithelial Morphogenesis. Developmental Cell. doi: 10.1016/j.devcel.2019.05.027.
– Proag A, Monier B, Suzanne M. (2019) Physical and functional cell-matrix uncoupling in a developing tissue under tension. Development. doi: 10.1242/dev.172577.
– Schott S, Ambrosini A, …, Suzanne M. (2017) A fluorescent toolkit for spatiotemporal tracking of apoptotic cells in living Drosophila tissues. Development. doi: 10.1242/dev.149807.
– Monier B, Gettings M, Gay G, Mangeat T, Schott S, Guarner A, Suzanne M. (2015) Apico-basal forces exerted by apoptotic cells drive epithelium folding. Nature. doi: 10.1038/nature14152.
Affiliation


