
Équipe
Responsables d’équipe : Devaud Jean-Marc & Isabel Guillaume
Présentation
Notre objectif est de comprendre la manière dont les réponses comportementales et les fonctions cérébrales sont influencées par différentes expériences de vie. Nous étudions, de manière intégrée, comment l’exposition aux interactions sociales (ex : compétition, coopération, reproduction) ou à d’autres facteurs environnementaux (ex : sources de nourriture, facteurs de stress) façonne les performances cognitives. Nous cherchons à élucider les bases nerveuses et génétiques de ces effets physiologiques ou pathologiques de l’expérience sur le comportement, et leurs interactions possibles avec l’âge (développement, vieillissement)
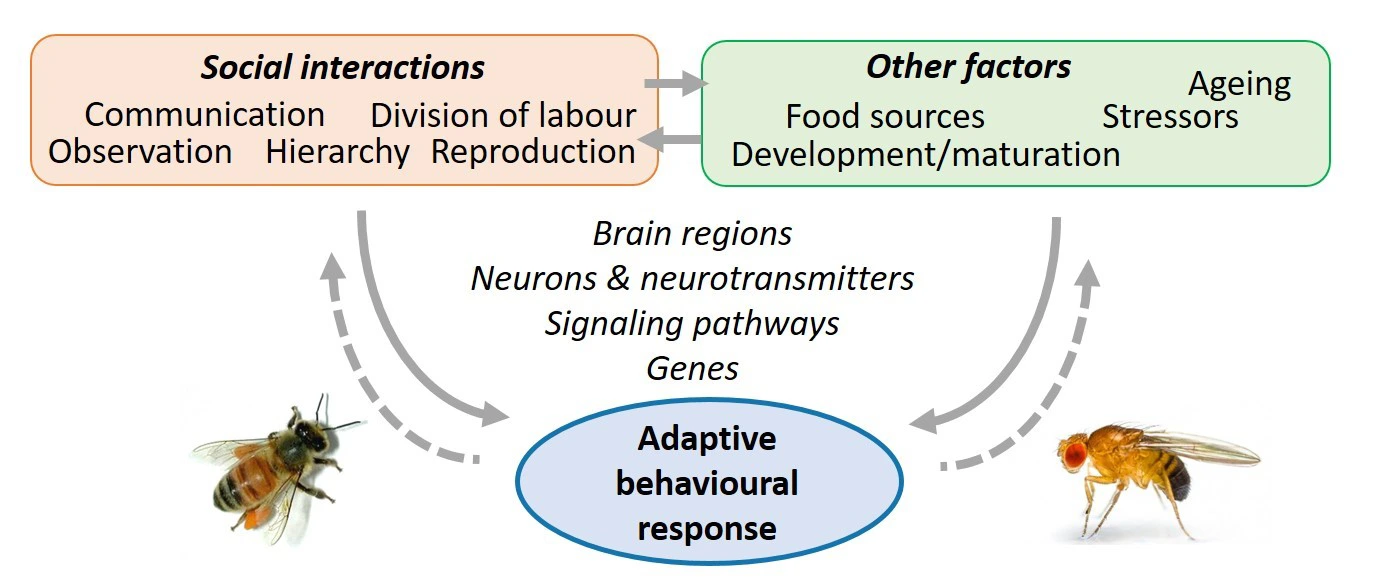
Ces questions sont abordées chez deux espèces d’insectes modèles : l’abeille domestique Apis mellifera et la drosophile Drosophila melanogaster, dont l’anatomie cérébrale et le génome sont bien décrits. Ce choix d’espèces présentant différents niveaux de complexité sociale nous permet d’intégrer l’étude de la plasticité dans un contexte social. Certains projets spécifiques impliquent également des comparaison avec d’autres espèces, y compris l’humain.
Notre approche est multidisciplinaire : neurosciences cognitives, génétique comportementale, neurobiologie des systèmes, psychologie expérimentale. Nous utilisons et développons des techniques de pointe d’observation comportementale et d’évaluation précise des performances cognitives, de neuroanatomie et d’imagerie fonctionnelle des régions cérébrales et circuits neuronaux sous-jacents, ainsi que des manipulations génétiques ou pharmacologiques ciblant les voies de signalisation impliquées.
Projet 1
Les ouvrières chez l’abeille mellifère subissent une maturation continue de leur cerveau et de leurs capacités d’apprentissage tout au long de leur vie, en lien avec leur aptitude à assumer différents rôles dans la colonie au fur et à mesure de leur vieillissement — une maturation neuronale et cognitive qui ne suit pas un programme de développement entièrement fixe. D’un côté, cette maturation peut être modulée par des signaux sociaux permettant d’adapter les comportements individuels aux besoins de la colonie. De l’autre, elle peut être perturbée si l’individu est exposé à un facteur de stress. Comment l’expérience individuelle des stimuli environnementaux ou sociaux influence-t-elle la maturation cérébrale, les capacités cognitives et la sensibilité au stress ?
Nous utilisons des tâches d’apprentissage olfactif impliquant la résolution d’ambiguïtés, telles que l’apprentissage de renversement, comme indicateur principal de la maturation cognitive et de la flexibilité comportementale. Nous avons déjà démontré que la capacité à réaliser un apprentissage de renversement nécessite une exposition précoce à l’environnement de la ruche, et qu’elle est modulée par l’expérience de butinage. Ces influences environnementales contribuent à façonner les circuits neuronaux dans certaines régions spécifiques du cerveau (les corps pédonculés), dont le fonctionnement est nécessaire pour ce type de tâche d’apprentissage complexe.
Nous abordons ainsi les questions suivantes :
– Quels facteurs environnementaux (par exemple, les substances odorantes présentes dans la ruche et/ou celles des fleurs, la lumière) influencent la maturation cérébrale et cognitive, et agissent-ils à des moments spécifiques de la vie adulte ?
– Comment une exposition chronique ou aiguë à des facteurs de stress module-t-elle le développement des corps pédonculés et/ou les capacités cognitives ? En particulier, comment la performance en apprentissage est-elle affectée par une exposition à des niveaux sublétaux de stress anthropiques (par exemple, les polluants) ?
– En retour, dans quelle mesure la capacité à répondre au stress varie-t-elle avec l’âge et/ou l’expérience individuelle ?
– Dans quelle mesure le statut social (par exemple, nourrice, butineuse…) ou la saison détermine-t-il la performance en apprentissage et la sensibilité au stress ?
– Les abeilles mellifères et les abeilles solitaires diffèrent-elles par leur niveau de flexibilité cognitive ?
– Quelles sont les populations neuronales et les circuits impliqués dans la régulation de ces réponses comportementales ?
– Quelles sont la dynamique et le niveau de plasticité des voies de signalisation identifiées comme régulateurs clés de la réponse au stress (par exemple, les allatostatines) et de l’apprentissage de renversement (par exemple, le GABA) ?
– Peut-on améliorer la résilience au stress chez l’abeille mellifère ?
Collaborations : R. Jeanson & C. Rampon (Toulouse), J. Carcaud & J.-C. Sandoz (Gif-sur-Yvette), M. Goubault & Ch. Lécureuil (Tours), M. Nouvian (Constance), C. Groh & W. Rössler (Wurtzbourg)
Projet 2
Comprendre comment les signaux sensoriels issus d’un environnement dynamique sont traités et traduits en comportements appropriés constitue une question centrale en neurosciences. L’agressivité, un comportement inné observé chez de nombreuses espèces, est essentielle pour l’acquisition de ressources, la défense de soi et l’établissement de hiérarchies sociales. Toutefois, elle est coûteuse sur le plan énergétique car elle implique une gamme d’actions allant des menaces aux confrontations physiques. Bien que l’agressivité soit généralement dépendante du contexte, une petite fraction d’individus présente une agressivité dysrégulée — cela concerne notamment 3 à 7 % des humains, entraînant d’importants problèmes sociétaux. L’agressivité est donc cruciale pour la survie animale, mais elle doit être finement régulée par des mécanismes inhibiteurs pour éviter toute escalade.
Notre projet vise à étudier les mécanismes fondamentaux et les circuits neuronaux qui permettent de réguler un comportement agressif approprié.
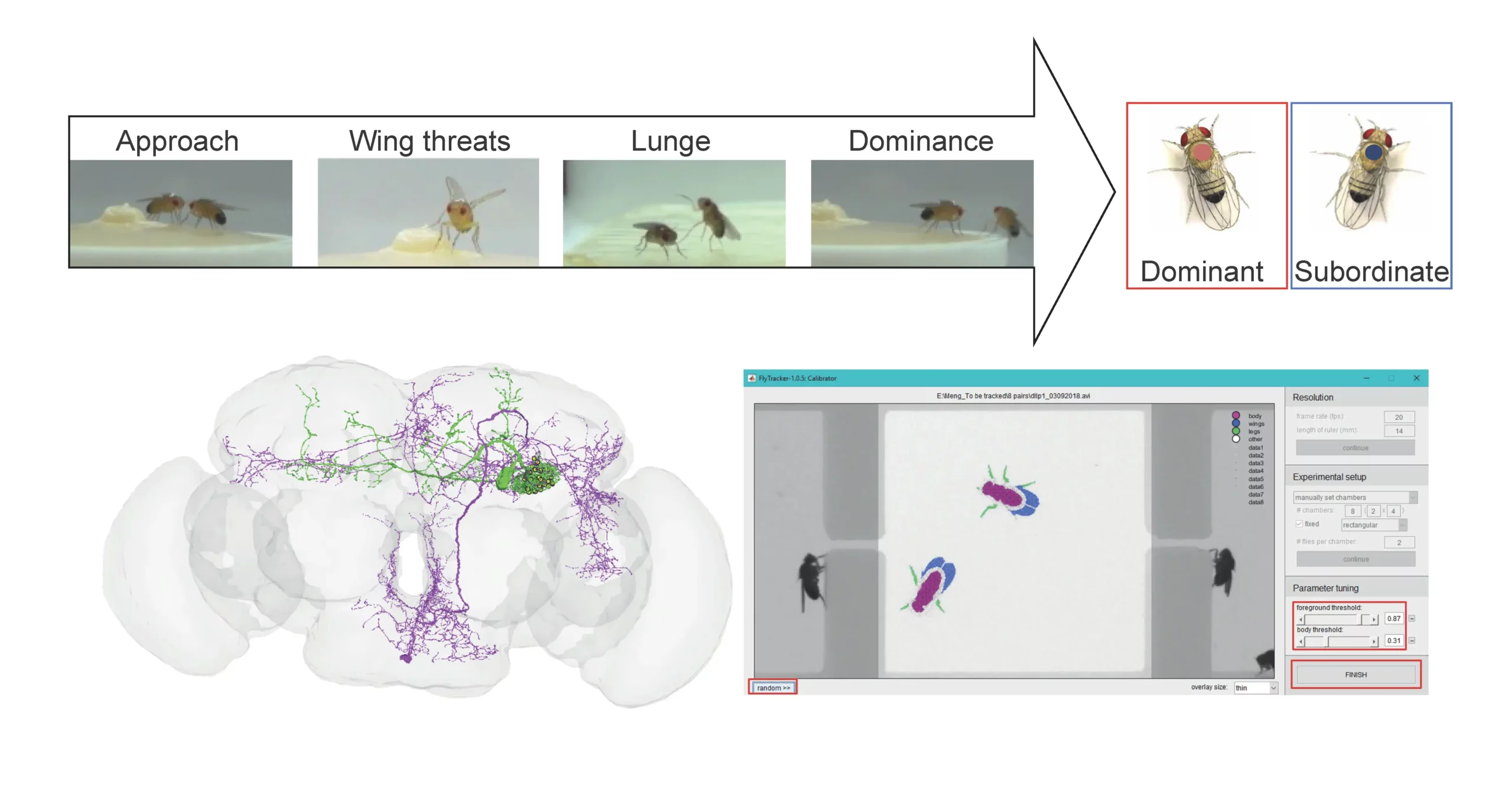
Pour aborder ces questions fondamentales, nous utilisons la mouche du vinaigre, Drosophila melanogaster, comme organisme modèle. Nous combinons des manipulations génétiques, différents tests comportementaux et de l’imagerie calcique in vivo pour répondre aux questions suivantes :
– Quels sont les circuits neuronaux dans le cerveau central qui régulent l’agressivité ?
– Quels sont les mécanismes fondamentaux sous-jacents ?
– Quels sont les signaux sensoriels essentiels pour déclencher des réponses agressives appropriées ?
– Comment ces signaux sont-ils transmis aux neurones moteurs pour générer des actions comportementales ?
– Dans quelle mesure les facteurs internes et externes influencent-ils l’expression de ce comportement ?
Projet 3
Le cerveau traite les informations sociales par le biais de deux types d’apprentissage : direct et indirect. L’apprentissage direct repose sur l’expérience personnelle, tandis que l’apprentissage indirect, tel que l’apprentissage par observation, consiste à apprendre en observant les autres. Le choix du partenaire est une décision cruciale qui a un impact direct sur la valeur sélective d’un individu. Ce choix peut être acquis par l’expérience personnelle ou par l’observation du comportement d’autrui. L’apprentissage par observation, ou mate-copying, permet aux individus de copier les choix des autres, ce qui peut entraîner une préférence pour des traits similaires.
En nous appuyant sur un paradigme expérimental bien établi (Figure X), nous chercherons à identifier et caractériser les neurones impliqués dans cet apprentissage social. Nous étudierons également comment le mate-copying peut influencer la structure des populations dans des conditions semi-naturelles. En combinant des approches neurobiologiques et écologiques, notre objectif est d’acquérir une compréhension globale de ce comportement d’apprentissage social.
Exemple typique d’une expérience de mate-copying chez la Drosophile melanogaster avec des mâles verts et roses
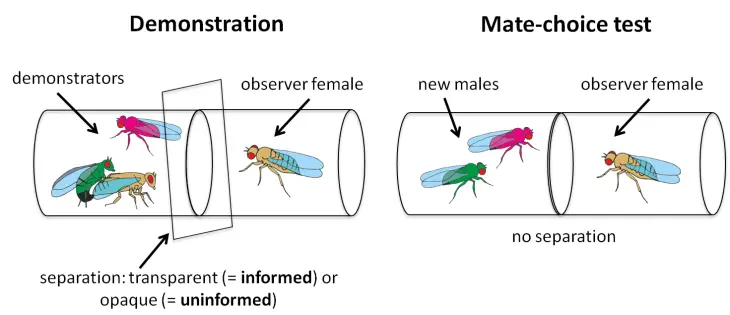
Après une phase de démonstration au cours de laquelle une femelle observatrice assiste à une copulation entre un mâle vert et une autre femelle, la femelle observatrice devra ensuite choisir entre un nouveau mâle vert et un mâle rose.
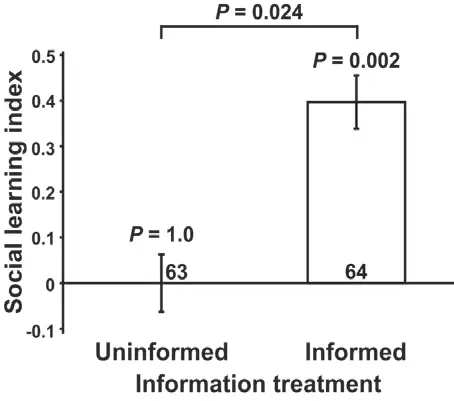
Ici, 0 % représente les cas où les deux phénotypes de mâles ont été choisis de manière aléatoire (c’est-à-dire dans 50 % des essais). La femelle observatrice aura tendance à choisir le mâle vert, c’est-à-dire le phénotype masculin choisi par la première femelle.
Projet 4
L’origine biologique ou culturelle de nos compétences mathématiques sophistiquées reste débattue. Bien que la plupart des caractéristiques de notre représentation des nombres soient partagées avec d’autres Vertébrés, il demeure difficile de trancher entre une faculté ancestrale héritée ou des mécanismes convergents. La distance évolutive avec les insectes offre une opportunité unique de comparer des systèmes numériques ayant évolué indépendamment.
En nous appuyant sur les premières preuves des compétences numériques chez les insectes, notre projet explore les mécanismes de traitement du nombre chez ces derniers, selon une approche comparative avec les Vertébrés. Nous visons à caractériser les propriétés du sens du nombre chez les insectes au niveau comportemental, en élaborant des protocoles cognitifs destinés à tester des caractéristiques clés du système des Vertébrés : par exemple, l’existence d’un double système pour les petites et les grandes quantités, ou encore une représentation du nombre intermodale.
Les zones cérébrales impliquées dans le sens du nombre chez les insectes seront identifiées à l’aide d’abeilles et de Drosophila, constituant une première étape décisive vers l’identification des corrélats neuronaux. Enfin, les données obtenues alimenteront des modèles computationnels permettant, pour la première fois, de concilier la modélisation de la sophistication cognitive des insectes avec les contraintes de perception, d’architecture et de connectivité cérébrales.
Le programme de recherche proposé apportera ainsi des éléments décisifs et opportuns pour une meilleure compréhension des contraintes physiologiques et écologiques qui permettent l’émergence des facultés cognitives numériques.

Figure : Une abeille doit choisir l’image contenant le plus petit nombre de formes afin d’obtenir une goutte de récompense sucrée (saccharose).
Membres de l'équipe
– Monchanin C, Drujont E, Le Roux G, Lösel PD, Barron AB, Devaud JM, Elger A, Lihoreau M (2024) Environmental exposure to metallic pollution impairs honey bee brain development and cognition. J Haz Mat 465, 133218 10.1016/j.jhazmat.2023.133218
– Sánchez-Morales A., Gigoux V, Matsoukas MT, Perez-Benito L, Fourmy D, Alibes R, Busqué F, Cordomí A, Devaud JM (2022) Reduction of stress responses in honey bees by synthetic ligands targeting an allatostatin receptor. Sci Rep 12, 167160 10.1038/s41598-022-20978-y
– Prunier A, Trannoy S (2024) Learning from fights: Males’ social dominance status impact reproductive success in Drosophila melanogaster. PLoS One 19(3):e0299839. 10.1371/journal.pone.0299839.
– Legros J, Tang G, Gautrais J, Fernandez MP, Trannoy S (2021) Long-term dietary restriction leads to development of alternative fighting strategies. (2021) Front Behav Neurosci 14;14:599676. 10.3389/fnbeh.2020.599676
– Finke V, Scheiner R, Giurfa M, Avarguès-Weber A (2023) Individual consistency in the learning abilities of honey bees: cognitive specialization within sensory and reinforcement modalities. Anim Cogn 26(3), 909-928. 10.1007/s10071-022-01741-2
– Avarguès-Weber A, Finke V, Nagy M, Szabó T, d’Amaro D, Dyer AG, Fiser J (2020) Different mechanisms underlie implicit visual statistical learning in honey bees and humans. Proc Natl Acad Sci 201919387. doi:10.1073/pnas.1919387117
– Muria A, Musso PY, Durrieu M, Portugal FR, Ronsin B, Gordon MD, Jeanson R, Isabel G (2021). Social facilitation of long-lasting memory is mediated by CO2 in Drosophila. Current Biology, 31(10) 2065-2074. doi.org/10.1016/j.cub.2021.02.044
– Nöbel S, Danchin E, Isabel G (2023) Mate copying requires the coincidence detector Rutabaga in the mushroom bodies of Drosophila melanogaster. i-Science, 26(9): 107682. doi.org/10.1016/j.isci.2023.107682
Affiliation

