
Équipe
Responsable d’équipe : Trouche Didier
Présentation
Les enzymes modifiant la chromatine et les modifications de la chromatine intègrent les signaux produits par les changements environnementaux extra ou intracellulaires afin d’adapter l’expression des gènes et d’établir la réponse cellulaire correcte. En outre, elles participent au maintien de l’information génétique au cours de la prolifération des cellules de mammifères. A ce titre, les enzymes modifiant la chromatine et les modifications de la chromatine jouent un rôle majeur dans des maladies telles que les cancers, qui sont caractérisés par un taux de prolifération accru et par une forte instabilité génétique.
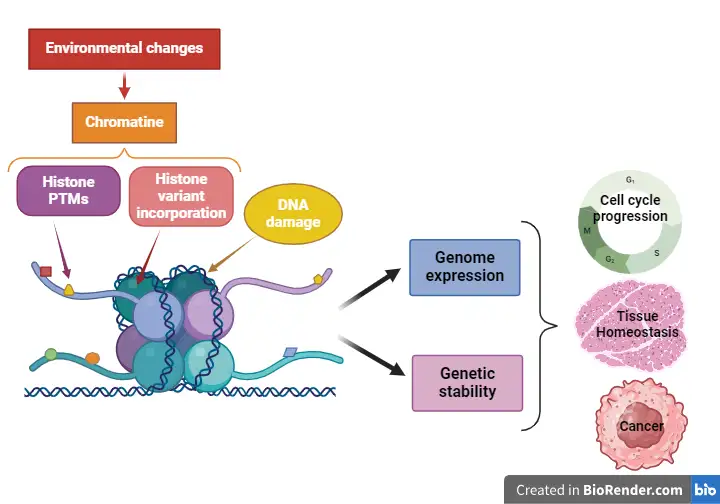
Notre équipe s’intéresse à la fonction des modifications de la chromatine dans la régulation de l’expression du génome et de la stabilité génétique, permettant ainsi le contrôle correct de processus tels que la progression du cycle cellulaire et le destin cellulaire. Plus spécifiquement, nous étudions la fonction du variant d’histone H2A.Z dans la transcription, le processing co-transcriptionnel de l’ARNm et la stabilité génétique au cours de la progression dans le cycle cellulaire. Nous nous intéressons également à la gestion des dommages oxydatifs de l’ADN dans le contexte de la chromatine et la façon dont ils affectent la structure et la fonction de la chromatine. Enfin, nous étudions la relation entre la structure de la chromatine et la vitesse de l’ARN pol II, en nous concentrant sur les mécanismes liés à l’élongation de la transcription, à la terminaison et au processing co-transcriptionnel de l’ARNm.
Projet 1
TROUCHE Didier, DR1 CNRS
CANITROT Yvan, DR2 CNRS
BERGOGLIO Valérie, CRCN CNRS
CHAILLEUX Catherine, IRCN CNRS
CINTORI Luana, Doctorante
Le thème général de nos travaux est l’étude des facteurs influençant les différents mécanismes de réparation de l’ADN et la stabilité génétique en réponse aux dommages causés à l’ADN. Notre approche se concentre sur le rôle de la chromatine et des facteurs qui affectent sa dynamique. Par l’utilisation et le développement d’outils cellulaires originaux, et en utilisant des approches cellulaires, moléculaires et d’imagerie ainsi que des données de génomique, nous tentons de disséquer la chorégraphie impliquée dans la gestion des dommages à l’ADN afin de mieux comprendre les conséquences de leurs défauts, dans le but de fournir de nouvelles cibles thérapeutiques pour améliorer l’efficacité des traitements qui ciblent l’ADN (cancers).
Nous nous concentrons actuellement sur les dommages oxydatifs de l’ADN, qui constituent la forme la plus courante de dommages à l’ADN. La façon dont ces dommages sont gérés dans les différents paysages chromatiniens et comment ils affectent la fonction chromatinienne sont encore mal compris, étant donné que la plupart des études ont utilisé des traitements oxydatifs qui induisent des dommages à l’ADN de façon aléatoire dans tout le génome. En outre, ces traitements entraînent de nombreux autres dommages oxydatifs que les dommages à l’ADN.
Nous avons développé des outils optogénétiques qui permettent la génération ciblée de dommages oxydatifs à l’ADN. Grâce à ce système original, nous étudions si des paysages chromatiniens spécifiques modulent de manière différentielle la gestion des dommages oxydatifs de l’ADN, et donc si certains loci génomiques sont plus enclins à la mutagenèse induite par les dommages oxydatifs. Nous étudions également les conséquences des dommages oxydatifs de l’ADN sur la fonction chromatinienne, c’est-à-dire la transcription et les processus liés à la transcription.
Références sélectionnées : Fages et al., 2020 ; Taty-Taty et al., 2016 ; Courilleau et al., 2012, Matterra et al., 2010.
Projet 2
TROUCHE Didier, DR1 CNRS
MUNIZ Lisa, CRCN CNRS
IACOVONE Marika, Doctorante
EISELE Maurane, Doctorante
Si l’information génétique est la même dans toutes les cellules de l’organisme, l’expression de cette information est spécifique de la fonction et du rôle de la cellule. La première étape de l’expression de l’information génétique, appelée transcription et réalisée par les ARN polymérases, consiste à lire l’information contenue dans un gène pour produire un ARN (Acide Ribonucléique). Les cellules de mammifères possèdent trois ARN polymérases différentes (appelées I, II et III) qui sont spécialisées dans la synthèse des différents ARN indispensables à la vie de la cellule. Les ARN messagers, qui codent pour les protéines, sont synthétisés par l’ARN polymérase II.
La transcription par l’ARN polymérase II a été largement étudiée, mais l’un des paramètres de ce processus, la vitesse d’élongation de l’ARN pol II (c’est-à-dire la vitesse à laquelle l’ARN polymérase se déplace sur l’ADN pour lire l’information contenue dans un gène, régulant ainsi le nombre de nucléotides ajoutés à l’ARN par unité de temps), reste mal caractérisé. On sait cependant que cette vitesse de transcription est très dynamique et extrêmement régulée à la fois au sein des gènes individuels et entre les gènes. De plus, la vitesse de transcription par l’ARN pol II est un déterminant majeur de l’identité des ARN produits à partir d’un gène. En effet, des études utilisant des mutants de l’ARN pol II transcrivant plus ou moins vite ont montré que la vitesse de transcription par l’ARN pol II peut contrôler l’épissage, la production d’ARN circulaires, la polyadénylation alternative et la terminaison de la transcription. Nous avons récemment montré qu’un changement localisé de la vitesse de transcription de l’ARN pol II se produit en aval de certains gènes en réponse à un stress oncogène, et que ce changement est corrélé à une augmentation de la transcription « read-through » en aval du gène. Nos travaux suggèrent donc que des changements localisés de la vitesse de transcription de l’ARN pol II lors de l’engagement dans un destin cellulaire particulier pourraient entraîner des changements dans le transcriptome. Cependant, on ne sait rien des facteurs qui régulent localement la vitesse de l’ARN pol II.
Nous avons construit un système rapporteur permettant de mesurer les changements de vitesse de l’ARN polymérase II. Nous avons utilisé ce système pour réaliser un criblage basé sur CRISPR/Cas9 afin d’identifier les facteurs régulant potentiellement la vitesse de l’ARN pol II. Nous sommes actuellement en train de caractériser la fonction des enzymes de modification de la chromatine issues de ce criblage dans la régulation de la vitesse de l’ARN pol II à des locus spécifiques du génome.
Références sélectionnées : Muniz et al., 2021a, Muniz et al., 2021b, Muniz et al., 2017
Projet 3
TROUCHE Didier, DR1 CNRS
FERNANDEZ-VIDAL Anne, MCUCN CNRS
ESCAFFIT Fabrice, CRHC CNRS
REYES Céline, IECN UT3-PS
IACOVONE Marika, Etudiante en thèse
Le nucléosome, composé d’un octamère de 4 histones autour duquel l’ADN est enroulé, est l’unité de base de la chromatine. Il peut subir des modifications qualitatives majeures telles que des modifications post-traductionnelles d’histones et/ou l’incorporation de variants d’histones à la place des histones canoniques à des endroits précis du génome, régulant ainsi les principales fonctions de la chromatine. Le variant d’histone H2A.Z est l’un des trois variants d’histone conservés de la levure à l’homme. Chez les vertébrés, H2A.Z est codé par deux gènes, donnant lieu à deux isoformes, H2A.Z.1 et H2A.Z.2, et à un variant d’épissage, H2A.Z.2.2.
H2A.Z est enrichi autour du site de démarrage de la transcription des gènes actifs et joue un rôle majeur dans le contrôle de la transcription de gènes spécifiques, régulant ainsi le destin cellulaire. H2A.Z semble également être nécessaire au maintien de l’information génétique. En effet, outre un rôle encore controversé dans la réparation des cassures double-brin de l’ADN, une fonction de H2A.Z dans la mitose et la réplication de l’ADN en phase S a été suggérée. Il est important de noter que les isoformes H2A.Z ou leurs régulateurs ont été trouvés surexprimés ou mutés dans de nombreux cancers.
Notre objectif est de caractériser la fonction des isoformes H2A.Z dans l’expression du génome, le contrôle du destin cellulaire et la stabilité génétique.
Pour surmonter les limitations sous-jacentes à l’utilisation de siRNAs, nous avons construit, par édition du génome, des lignées cellulaires nous permettant d’induire la dégradation des isoformes H2A.Z endogènes de manière rapide, inductible et réversible. Nous avons utilisé ces lignées cellulaires pour effectuer un criblage génomique CRIPR/Cas9 afin d’identifier les gènes synthétiquement létaux avec les isoformes H2A.Z, ce qui a conduit à l’identification de nombreuses voies pertinentes. Nous avons également généré des lignées cellulaires pour suivre la localisation de H2A.Z endogène en temps réel et des lignées cellulaires avec différentes capacités à s’engager dans différents destins. Nous utilisons maintenant ces lignées cellulaires pour caractériser la fonction directe des isoformes H2A.Z dans la transcription, la réparation de l’ADN lors de la progression dans le cycle cellulaire et le contrôle du destin cellulaire.
Références sélectionnées : Rispal et al., 2024, Lamaa et al., 2020, Rispal et al., 2019, Lazorthes et al., 2015
Membres de l'équipe
– Rispal J, Rives C, Jouffret V, Leoni C, Dubois L, Chevillard-Briet M, Trouche D, Escaffit F. Control of intestinal stemness and cell lineage by histone variant H2A.Z isoforms (2024) Mol Cell Biol. 18:1-18.
– Ouvrard J, Muniz L, Nicolas E, Trouche D. (2022) Small Interfering RNAs Targeting a Chromatin-Associated RNA Induce Its Transcriptional Silencing in Human Cells. Mol Cell Biol. 42(12):e0027122
– Muniz L, Nicolas E, Trouche D. (2021) RNA pol II speed: a key player in control and adaptation of transcriptome composition, EMBO J. 40(15): e105740.
– Muniz L, Lazorthes S, Delmas M, Ouvrard J, Aguirrebengoa M, *Trouche D, *Nicolas E (* co-last authors) (2021). Circular AnRIL RNAs switch from repressor to activator of p15/CDKN2B expression during RAF1 oncogene-induced senescence. RNA Biol. 10.1080.
– Fages J, Chailleux C, Humbert J, Jang S-M, Loehr J, Lambert J-P, Coté J, *Trouche D, *Canitrot Y (* co-last authors). (2020) JMJD6 participates in the maintenance of ribosomal DNA integrity in response to DNA damage. PLoS Genetics, 16(6): e1008511.
– Lamaa A, Humbert J, Aguirrebengoa M, Xue C, Nicolas E, Coté J, Trouche D. (2020) Integrated analysis of H2A.Z isoforms functions reveals a complex interplay in gene regulation. eLife 28 (9), doi: 10.7554/eLife.53375.
– Rispal J, Baron L, Beaulieu J-F, Chevillard-Briet M, *Trouche D, *Escaffit F (*co-last authors) (2019) The H2A.Z histone variant integrates Wnt signaling in intestinal epithelial homeostasis. Nature Comm 10 (1): 1827.
– §Muniz L., §Deb M.K. (§co-first authors), Aguirrebengoa M, , Lazorthes S., *Trouche D, *Nicolas E. (* co-last authors) (2017) Control of gene expression in senescence through transcriptional read-through of convergent protein-coding genes. Cell Reports 21(9):2433-2446.
– Grezy, A., Chevillard-Briet, M., *Trouche, D., *Escaffit, F. (* co-last authors) (2016) Control of genetic stability by a new heterochromatin compaction pathway involving the Tip60 histone acetyltransferase. Mol Biol Cell 15: 599-607.
– Taty-Taty, G., Chailleux, C., Quaranta, M., So, A., Guirouilh-Barbat, J., Lopez, B., Bertrand, P., Trouche, D., Canitrot, Y. (2016) Control of alternative end joining by the chromatin remodeler p400 ATPase. Nucleic Acids Res 44: 1657-1668.
– Salifou, K, Ray, S, Verrier, L., Aguirrebengoa, M., Trouche, D., *Panov, K.I., *Vandromme, M. (* co-last authors) (2016) The histone demethylase JMJD2A/KDM4A links nutrients/growth factors availability and ribosomal RNA transcription. Nature Comm. 7: 10174.
– Lazorthes, S., Vallot, C. Briois, S., Aguirrebengoa, M, Thuret, J.-Y., St. Laurent, G., Rougeulle, C., Kapranov, P., Mann, P., *Trouche, D, *Nicolas, E (* co-last authors). (2015) A vlincRNA participates in senescence maintenance by relieving H2AZ-mediated repression at the INK4 locus in trans, Nature Comm. 6:5971.
– Chevillard-Briet, M., Quaranta, M., Grezy, A., Matterra, L., Courilleau, C., Philippe, M., Mercier, P., Corpet, D., Lough, J., Ueda, T., Fukunaga, R., *Trouche, D., *Escaffit, F. (*co-last authors). (2014) Interplay between chromatin modifying enzymes controls colon cancer progression through Wnt signaling. Human Mol Genet 23: 2120-31.
– Courilleau, C., Chailleux, C., Jauneau, A., Grimmal, F., Briois, S., Boutet-Robinet, E., Boudsocq, F., *Trouche, D., *Canitrot, Y. (*co last-authors) (2012) The chromatin remodeler p400 ATPase facilitates Rad51 mediated repair of DNA double-strand breaks. J Cell Biol, 199: 1067-1081.
– Verrier, L., Escaffit, F., Chailleux, C., *Trouche, D., *Vandromme, M. (* co-last-authors). (2011) A new isoform of histone demethylase JMJD2A/KDM4A required for skeletal muscle differentiation. Plos Genet. 2011 7(6):e1001390.
– Capp, J.P., Boudsocq, F., Bergoglio, V., Trouche, D., Cazaux, C., Blanco, L., Hoffmann, J.S., Canitrot, Y. (2010) The R438W polymorphism of human DNA polymerase lambda triggers sensitivity to camptothecin by compromising the homologous recombination pathway. Carcinogenesis, 31(10): 1742-7.
– *Mattera, L., *Courilleau, C. (*co-first authors), Legube, G., Ueda, T., Fukunaga, R., Chevillard-Briet, M., §Canitrot, Y., §Escaffit, F. (§ equal contribution), Trouche, D. (2010) The E1A-associated p400 protein modulates cell fate decision by the regulation of ROS homeostasis. PLos Genet. 6(6):e1000983.
– *Chailleux, C., *Tyteca, S. (*co-first authors), Courilleau, C., Papin, C., Boudsocq, F., Puget, N., Grigoriev, M., Canitrot, Y., Trouche, D. (2010) Physical interaction between the histone acetyl transferase Tip60 and the DNA double strand breaks sensor MRN complex. Biochem J., 426: 365-371.
– *Mattera, L., *Escaffit, F. (*co-first authors), Pillaire, M.-J., Selves, J., Tyteca, S., Hoffmann, J.-S., Gourraud, P.-A., §Chevillard-Briet, M., §Cazaux, C. (§ equal contribution), Trouche, D. (2009) The p400/Tip60 ratio is critical for colorectal cancer cells proliferation through DNA damage response pathways. Oncogene, 28: 1506-1517.
– Tyteca, S., Vandromme, M., Legube, G., Chevillard-Briet, M., Trouche, D. (2006) Tip60 and p400 are both required for UV-induced apoptosis but play antagonistic roles in cell cycle progression. EMBO J., 25: 1680-1689. Selected by EMBO J editors as one of the four best papers published by EMBO J during the first half of 2006 (EMBOencounters 6, summer 2006)
– *Daury, L., *Chailleux, C. (*co-first authors), Trouche, D. (2006) Deposition of histone H3.3 on E2F responsive genes is linked to transcription. EMBO R. 7: 66-71. Régnier, V. (corresponding author), Vagnarelli, P., Fukagawa, T., Zerjal, T., Burns, E., Trouche, D., Earnshaw, W., Brown, W. R. (2005) CENP-A is required for accurate chromosome segregation and sustained kinetochore association of BubR1. Mol. Cell. Biol. 25: 3967-3981.
– Legube, G., *Linares, L.K., *Tyteca, S. (*co-second athors), Caron, C., Scheffner, M., Chevillard-Briet, M., and Trouche, D. (2004) Role of the histone acetyl transferase Tip60 in the p53 pathway. J. Biol. Chem. 279: 44825-44833.
– Legube, G., Linares, L.K., Lemercier, C., Scheffner, M., Khochbin, S., and Trouche, D. (2002) The histone acetyl transferase Tip60 is targeted to proteasome-mediated degradation by the Mdm2 oncoprotein. EMBO J. 21:1704-1712.
Affiliation




