
Équipe
Responsable d’équipe : Cuvier Olivier
Présentation
Nous étudions comment la dynamique de la chromatine régit le destin et la prolifération des cellules souches par le biais de :
a) le contrôle de l’accessibilité des caractéristiques génomiques clés telles que les promoteurs de gènes, les corps de gènes ou les enhancers, notamment par le positionnement des nucléosomes.
b) le repliement en 3D de la chromatine qui joue un rôle clé dans la régulation de la transcription, de la réplication et de la réparation de l’ADN, ce qui implique un regroupement en 3D ou des hubs et des boucles d’ADN entre des éléments fonctionnels distincts.
c) la régulation des ARN non codants (ARNnc) par des cofacteurs spécifiques tels que les ARN hélicases.
d) la démarcation épigénétique par les barrières chromatiniennes et les modifications histoniques couplées à la transcription – les histones méthyltransférases de H3K36 (dNSD/Mes-4, dSet2/Hypb, etc.), qui participent à la régulation des oncogènes en fonction du positionnement des nucléosomes et des plaques tournantes de la chromatine.
À cette fin, nous utilisons la biochimie, la génétique moléculaire, la biologie cellulaire combinée à la biologie computationnelle des données de séquençage à haut débit pour aborder les aspects clés de l’organisation de la chromatine.
Projet 1
Les interactions entre promoteurs et activateurs mettent en relation des activateurs distants et des promoteurs cibles, ce qui implique des contacts dynamiques à longue portée. Nous avons développé des schémas de purification biochimique des complexes insulateurs de la chromatine permettant d’identifier les cofacteurs des protéines insulatrices, y compris les facteurs participant à la formation de clusters 3D, impliquant CP190, Cohesin, et Chromator. Nous avons également trouvé des régulateurs épigénétiques tels que NSD/dMes-4 qui participent à la définition de hubs en 3D, similaires aux îlots de chromatine dans les cellules tumorales. Enfin, nous avons identifié de nouveaux cofacteurs qui régulent la stabilité des ARN non codants, y compris les hélicases ARN.
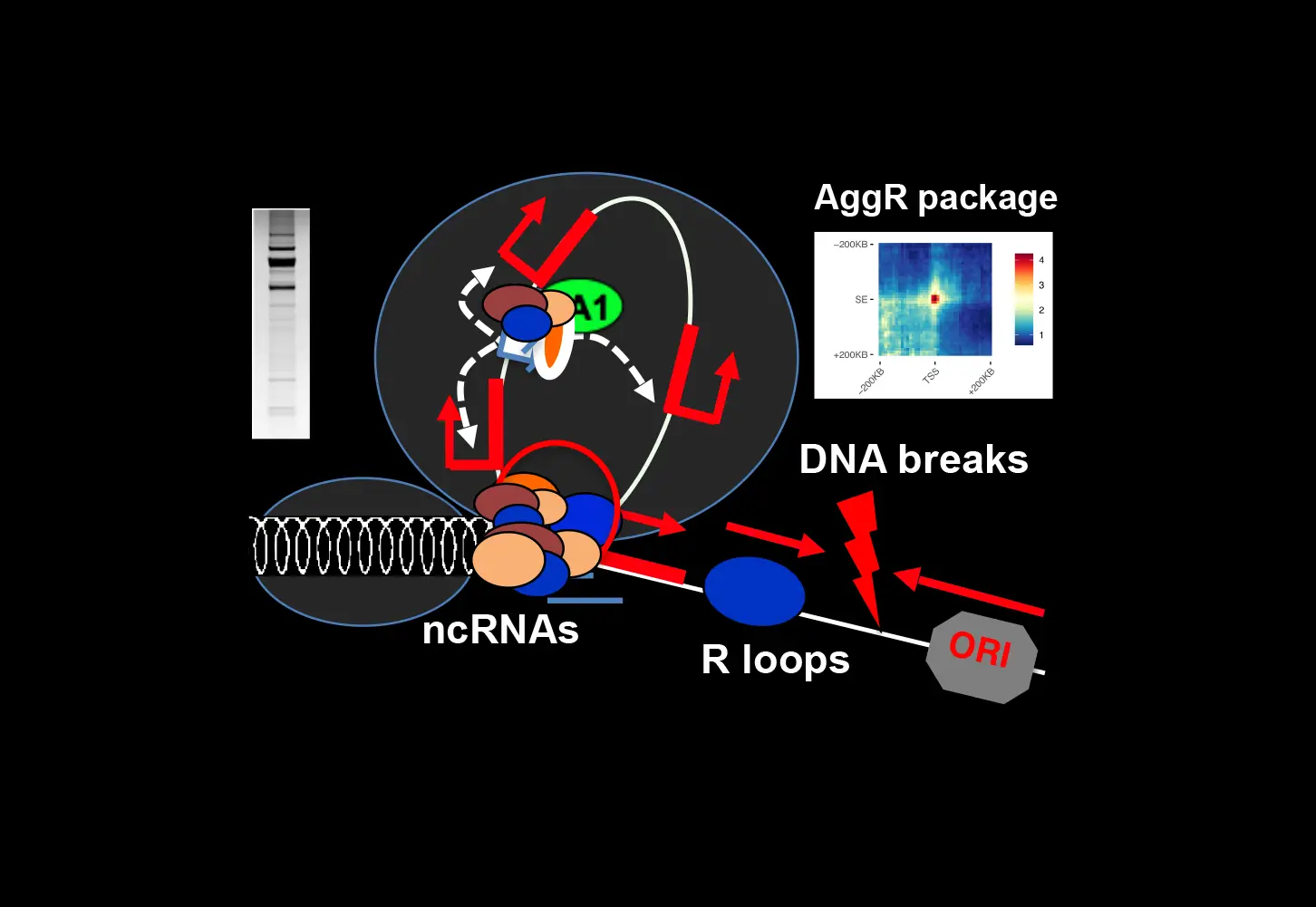
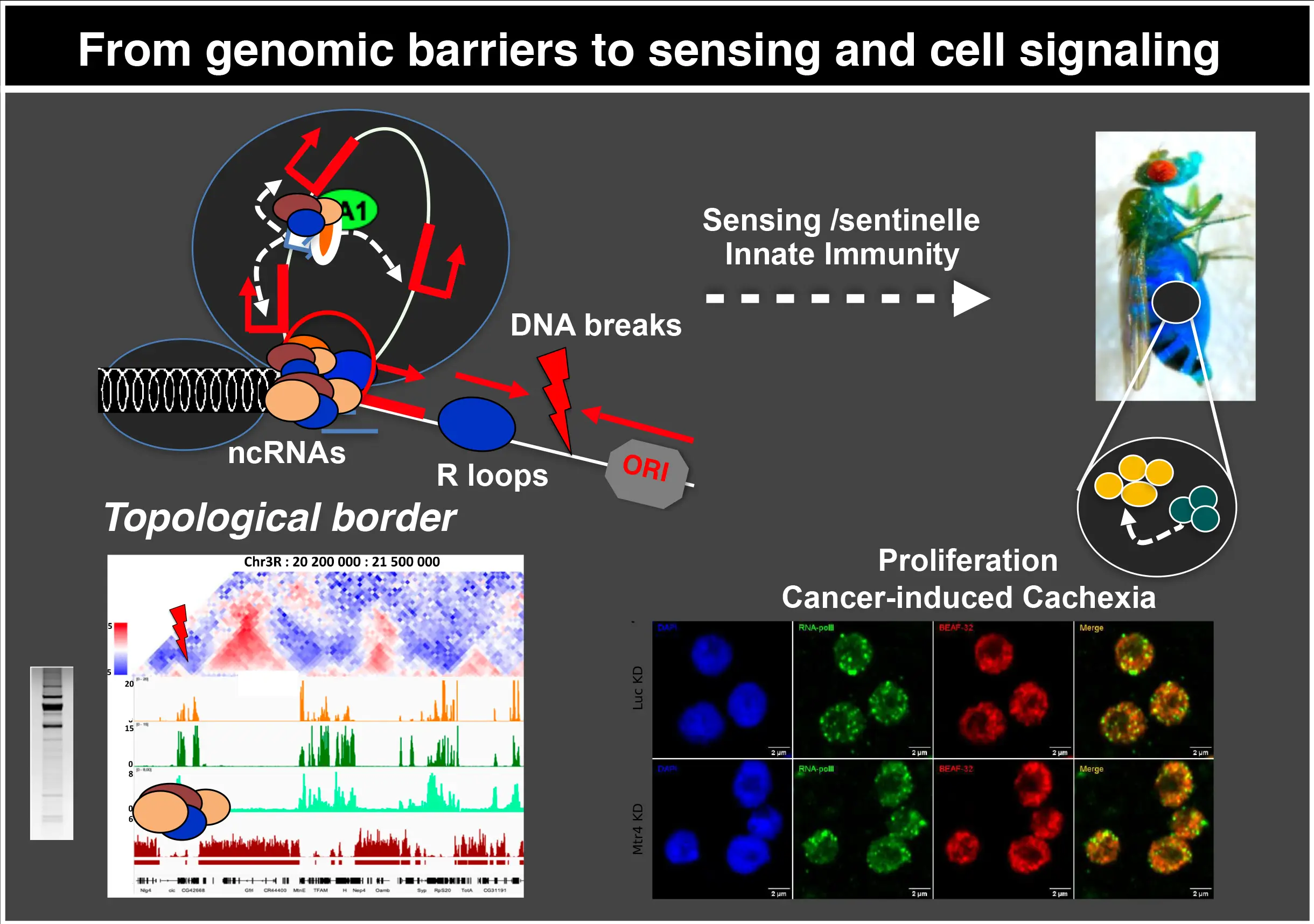
En parallèle, nous développons des approches de biologie informatique, notamment la méthode intitulée Agrégation, qui permet de détecter les contacts à longue portée même s’ils sont moins stables que ceux formés aux frontières du TAD, par exemple sous forme de boucles d’isolateur, et nous avons maintenant mis à disposition un package de ressources (Bioconductor package : HiCAggR). L’agrégation permet de sonder les contacts à longue portée à très haute résolution (< 1 kb) et donc d’identifier les cibles génétiques appropriées.
En outre, nous pouvons maintenant aborder systématiquement la manière dont les contacts transitoires à longue portée peuvent entraver la dérégulation fortuite des gènes au-delà des frontières des TAD ou à travers les isolateurs en utilisant et en intégrant les données de nos approches omiques, microscopiques et CRISPR à l’échelle de la cellule unique. Notamment, les contacts transitoires enhancer-promoteur sont sous l’influence de facteurs tels que le cohesin qui peut médier l’extrusion des boucles, les isolateurs et les ARN non codants. L’accumulation d’ARNnc par plusieurs moyens, y compris la déplétion de l’ARN hélicase, peut entraver la formation de boucles 3D en régulant l’attachement ou la stabilité des protéines insulatrices et des cofacteurs contenant un domaine de liaison à l’ARN. Grâce à un grand nombre de données préliminaires, nous étudions ces mécanismes en combinant des approches d’ingénierie génomique CRIPR/dCas9 et des approches computationnelles. Nous analysons comment l’accumulation d’ARNnc régule la dynamique des insulateurs et des cofacteurs et les contacts à longue distance associés entre l’enhancer et les promoteurs cibles distants, établissant un lien entre les régulations structurelles et fonctionnelles.
Projet 2
L’immunité innée est une forme ancienne de défense de l’hôte qui a été conservée tout au long de l’évolution. Comme les mammifères, les mouches codent un certain nombre de récepteurs de reconnaissance des formes (PRR) qui reconnaissent des motifs pathogènes conservés appelés motifs moléculaires associés aux pathogènes (PAMP). En réponse à une infection microbienne et suite à l’activation des voies Toll ou IMD, les cellules du corps gras produisent des peptides antimicrobiens qui sont libérés dans l’hémolymphe. Grâce à la voie Sting, la drosophile est également capable de reconnaître les dinucléotides cycliques, une autre classe de PAMP. Cela lui permet de reconnaître à la fois les bactéries et les virus, ce qui entraîne l’activation de Relish, l’homologue NF-κB de la drosophile, et la production de peptides antimicrobiens. À ce jour, bien que la voie cGAS/Sting soit impliquée dans la reconnaissance cytosolique de fragments d’ADN ou d’hybrides ADN/ARN provenant de cassures de l’ADN chez l’homme, et que cGLR1, un orthologue de cGAS, soit capable de détecter l’ARNdb pour induire une réponse immunitaire via dSTING, la fonction de cette voie dans la détection des dommages à l’ADN n’a pas encore été étudiée chez la drosophile. Après avoir caractérisé la présence de cassures de l’ADN et/ou de boucles R dans plusieurs contextes tumorigènes, nous étudions la fonction de la voie cGAS/Sting, d’abord dans la détection des dommages à l’ADN et ensuite dans son implication dans la progression tumorale.
Chez l’homme : En collaboration avec le groupe de M. Benkirane, nous avons cherché à décrypter la fonction nucléaire de cGAS/Sting chez l’homme. cGAS/Sting joue un role central dans la signalisation de l’ADN viral étranger ou dans l’ADN illégitime pour déclencher une réponse cellulaire appropriée vers l’immunité innée. Nous avons effectué des analyses bioinformatiques de ces données, en revisitant les dimensions chromatiniennes de la signalisation de cGAS depuis les sites endogènes jusqu’à l’induction d’IRF3 et de la voie de l’immunité innée associée. Notre travail vise à comprendre comment les génomes hôtes reconnaissent structurellement et fonctionnellement l’ADN étranger et interagissent éventuellement avec les virus, ou pour une lutte efficace par des moyens immunologiques.
Chez la drosophile, nous étudions ces voies pour caractériser la détection endogène de l’ARN-ADN en cas de stress oncogène, de défauts de pause, ou lors de la production non contrôlée d’ARNnc et de boucles R, dans le contexte des hélicases ARN identifiées dans notre équipe, qui servent par ailleurs à traiter l’ADN viral. Nous caractérisons l’impact de leurs dysfonctionnements sur les cellules souches adultes en transformation, afin de mieux comprendre comment la voie de l’immunité innée protège le génome dans un tel contexte.
Projet 3
Notre expertise en biologie computationnelle a été intégrée dans un réseau de collaborations avec des équipes travaillant sur les virus (laboratoires de M Benkirane ; R. Kiernan ; C. Neuveut ; D. Durantel ; S. Emiliani ; E. Bertrand, etc.) ainsi que dans le développement d’un nouveau GDR, le réseau DynaVIR. Notre objectif est de déchiffrer si et comment la dynamique de la chromatine, soit par l’accessibilité locale de la chromatine, soit plus globalement par des interactions 3D, peut réguler l’intégration et/ou la réactivation virale, par exemple par le repositionnement local des nucléosomes ou par des boucles 3D, par exemple lorsque le génome viral interagit avec les gènes voisins. Nous avons abordé les mécanismes impliquant la latence virale et/ou la (ré)activation qui sont cruciaux pour les cellules cancéreuses induites par le virus, comme dans l’hépatocarcinome (Collab : D. Durantel, C. Neuveut). Nous analysons comment l’intégration virale du VHB agit sur les interactions chromatiniennes locales, sur les marques épigénétiques, ou sur les repliements d’ordre supérieur à l’échelle des clusters 3D, des TADs ou des compartiments au sein des cellules infectées de patients. Nous évaluons les changements à l’échelle des contacts à longue portée, entre le génome viral et le paysage chromatinien voisin, y compris les oncogènes, dans des contextes cellulaires spécifiques et de développement de cancer ou non, qui peuvent être éventuellement testés par des approches dCas9/CRAB cherchant à démêler les liens de causalité entre l’intégration du virus et l’induction d’oncogènes (Collab : Diogo-Dias, Neuveut, Durantel).
– Heurteau A, Perrois C, Depierre D, Fosseprez O, Humbert J, Schaak S, Cuvier O#. Insulator-based loops mediate the spreading of H3K27me3 over distant micro-domains repressing euchromatin genes. (2020) Genome Biol. doi: 10.1186/s13059-020-02106-z.PMID: 32746892
– Machida S, Depierre D*, Chen HC, Thenin-Houssier S, Petitjean G, Doyen CM, Takak M, Cuvier O#, Benkirane M*. Exploring histone loading on HIV DNA reveals a dynamic nucleosome positioning between unintegrated and integrated viral genome. (2020). Proc Natl Acad Sci U S A. doi: 10.1073/pnas.1913754117.
– Salifou K*, Burnard C*, Basavarajaiah P, Grasso G, Helsmoortel M, Mac V, Depierre D, Franckhauser C, Beyne E, Contreras X, Dejardin J, Rouquier S, Cuvier O, Kiernan R. Chromatin-associated MRN complex protects highly transcribing genes from genomic instability. (2021) Sci Adv. doi: 10.1126/sciadv.abb2947.
– Thenin-Houssier S, Machida S, Jahan C, Bonnet-Madin L, Abbou S, Chen HC, Tesfaye R, Cuvier O, Benkirane M. POLE3 is a repressor of unintegrated HIV-1 DNA required for efficient virus integration and escape from innate immune sensing. (2023) Sci Adv. doi: 10.1126/SciAdv.adh3642.
– Recoules L, Heurteau A, Raynal F, Karasu N, Moutahir F, Bejjani F, Jariel-Encontre I, Cuvier O, Sexton T, Lavigne AC, Bystricky K. The histone variant macroH2A1.1 regulates RNA polymerase II-paused genes within defined interaction landscapes. (2022) J Cell Sci. doi: 10.1242/jcs.259456.
– Depierre D, Perrois C, Schickele N, Lhoumaud P, Abdi-Galab M, Fosseprez O, Heurteau A, Margueron R, Cuvier O#. Chromatin in 3D distinguishes dMes-4/NSD and Hypb/dSet2 in protecting genes from H3K27me3 silencing. (2023) Life Sci Alliance. doi:10.26508/lsa.202302038.
– Contreras X*, Depierre D*, Akkawi C, Srbic M, Helsmoortel M, Nogaret M, LeHars M, Salifou K, Heurteau A, Cuvier O, Kiernan R. PAPγ associates with PAXT nuclear exosome to control the abundance of PROMPT ncRNAs. Nat Commun. (2023). doi:10.1038/s41467-023-42620-9.
– Mancheno-Ferris A, Immarigeon C, Rivero A, Depierre D, Schickele N, Fosseprez O, Chanard N, Aughey G, Lhoumaud P, Anglade J, Southall T, Plaza S, , Payre F, Cuvier O#, Polesello C#. Crosstalk between chromatin and Shavenbaby defines transcriptional output along the Drosophila intestinal stem cell lineage. (2023). iScience. doi: 10.1016/j.isci.2023.108624.
– Schickele, N Tesfaye, Schaak, Martin P. and O. Cuvier. HiCAggR –Bioconductor package https://www.bioconductor.org/packages/release/bioc/html/HicAggR.html
– Fosseprez O, Cuvier O Uncovering the functions and mechanisms of regulatory elements-associated non-coding RNAs. Biochim Biophys Acta Gene Regul Mech. (2024). doi:10.1016/j.bbagrm.2024. 195059.
Affiliation




