
Équipe
Responsable d’équipe : Wang Xiaobo
Présentation
La migration cellulaire est cruciale pour le développement et le cancer. Les cellules migratrices présentent différents comportements migratoires et réponses chimio-mécaniques lorsqu’elles rencontrent différents microenvironnements, comme la matrice et les cellules substrats. Ainsi, une question importante est de savoir comment les cellules en migration interagissent avec différents microenvironnements et y répondent. De plus, la migration cellulaire est souvent intégrée à d’autres processus cellulaires tels que la prolifération cellulaire. La manière dont la migration cellulaire s’intègre à la prolifération cellulaire est essentielle pour comprendre l’initiation et la progression du cancer.
L’équipe utilise un ensemble complémentaire de stratégies moléculaires, génétiques et d’imagerie ainsi que de modélisation mathématique et physique pour étudier 1) comment différents types de migration cellulaire se produisent, sont contrôlés et jouent un rôle dans la morphogenèse des tissus et la progression du cancer, et 2) comment la migration cellulaire est liée et intégrée à la division cellulaire au cours de la progression du cancer. L’équipe est expérimentée dans de nombreuses techniques d’imagerie (biosenseurs FRET, optogénétique, microscopie super-résolution RIM et microscopie à expansion) et certains outils de biophysique (ablation au laser, pince optique).
Projet 1
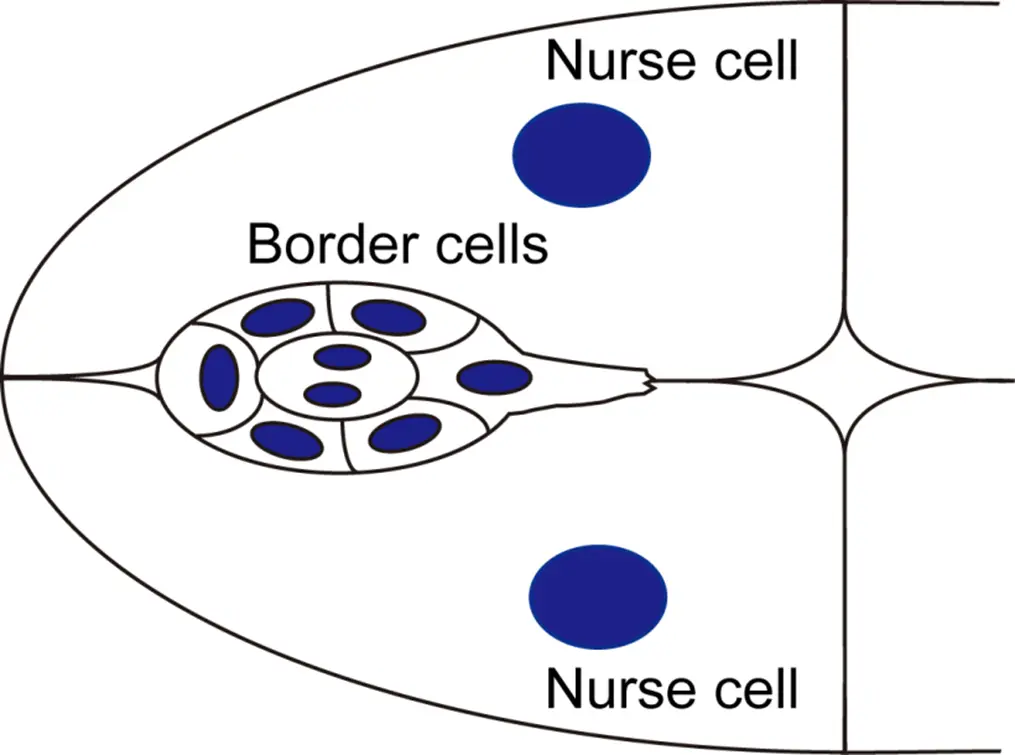
La migration cellulaire collective au cours du développement et des processus pathologiques se produit souvent dans les tissus riches en cellules, composés non seulement de matrices mais également de cellules substrats. Les caractéristiques et le rôle des cellules substrats sont mal compris par rapport à ceux de la matrice. L’équipe utilise la migration des cellules de bordure de l’ovaire de drosophile comme modèle in vivo pour comprendre comment les cellules collectives interagissent et communiquent avec les cellules substrats pour traverser un tissu riche en cellules.
Notre étude précédente a identifié les impulsions d’actomyosine comme un facteur critique contrôlant la migration des cellules de bordures (Combedazou et al, J Cell Sci, 2017). Nous avons également révélé deux pools fonctionnels de Rac1 au niveau des protusions et des câbles des cellules de bordures, et l’équilibre entre ces deux pools de Rac1 permet d’intégrer la direction et la coordination de la migration des cellules de bordures (Zhou et al, Nat Commun, 2022). Concernant les cellules de bordures, nous explorerons l’origine, le contrôle et le rôle des impulsions d’actomyosine dans l’efficacité de la migration. De plus, nous avons récemment révélé que Rac1 est également essentiel aux cellules substrat pour le trajet de déplacement des cellules migratoires, affectant ainsi l’efficacité de la migration. Concernant les cellules substrats, nous explorerons les facteurs clés présents dans les cellules substrats qui contrôlent le comportement et l’efficacité de la migration des cellules de bordures, ce qui est peu exploré. Comprendre l’environnement constitué de cellules substrats nous permettra de mieux comprendre comment les cellules cancéreuses qui migrent à travers un environnement cellulaire similaire au stroma lors de l’invasion du cancer et des métastases.
Projet 2
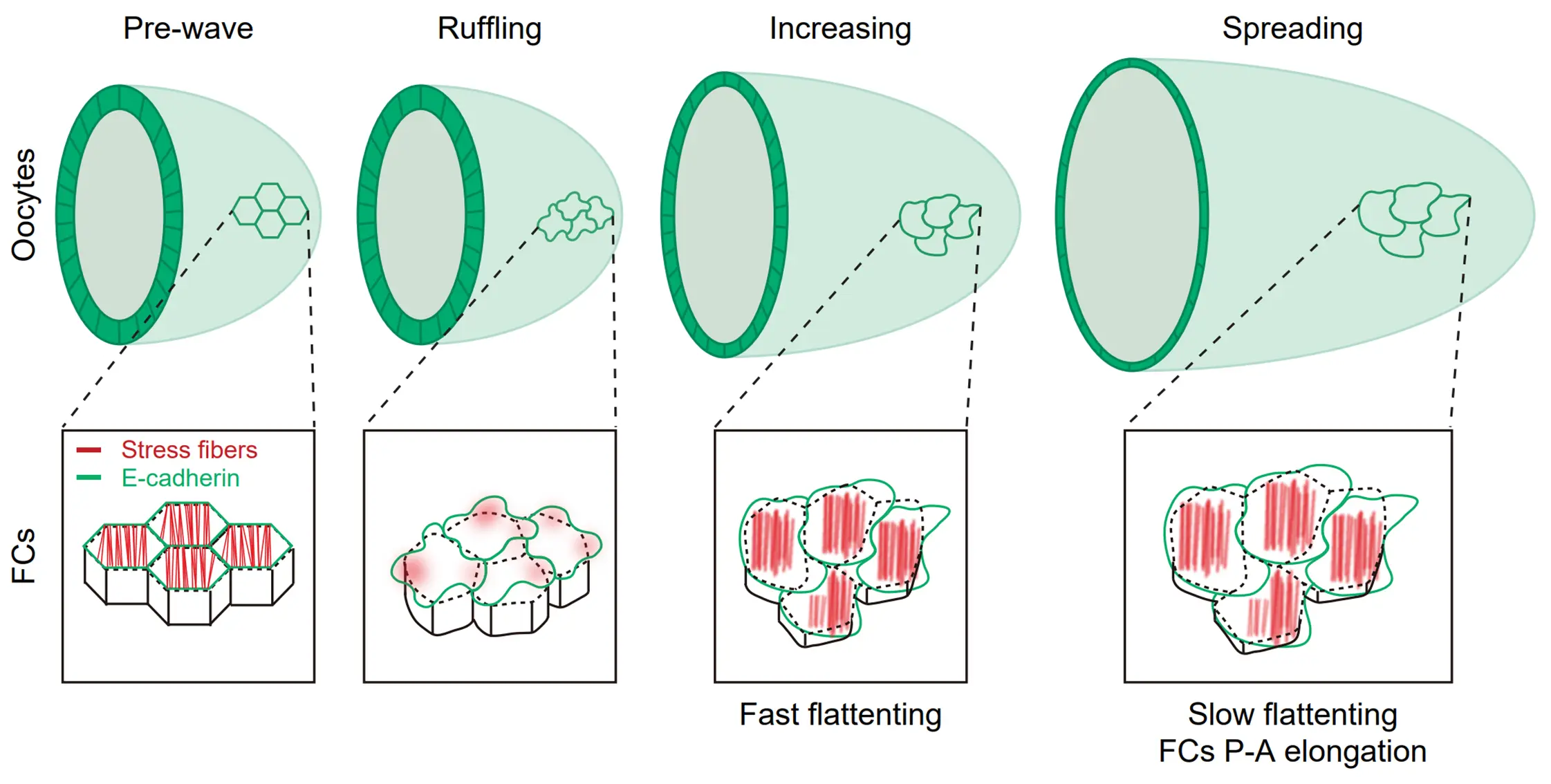
Les fibres de stress constituent l’un des importants réseaux du cytosquelette par lequel les cellules épithéliales interagissent et communiquent avec la matrice extracellulaire et jouent ainsi divers rôles dans le contrôle de la prolifération, de la migration et de l’homéostasie cellulaire. Par rapport aux cellules en culture, notre compréhension des fibres de stress in vivo est très limitée. L’équipe utilise les cellules épithéliales folliculaires de drosophile comme modèle in vivo pour comprendre l’origine, la modulation, la mécanique et la fonction des fibres de stress basales dans les cellules épithéliales in vivo.
Notre étude précédente a fourni des informations mécanistiques sur les fibres de stress basale contractiles : 1) interaction avec différentes adhérences (Qin et al, Nat Commun, 2017) ; 2) mécanisme de contrôle des impulsions basales de myosine (Qin et al, Nat Commun, 2018) ; 3) réseau de cytosquelette supracellulaire (Popkova et al, Nat Commun, 2020).
Concernant ces fibres de stress contractile, nous explorerons deux questions principales : 1) le contrôle des réseaux et impulsions basales de F-actine, et 2) l’origine et la propagation des impulsions basales d’actomyosine. De plus, nous avons récemment découvert des fibres de stress basales en expansion qui contrôlent un nouveau comportement d’expansion épithéliale pour intégrer l’aplatissement des cellules et l’élongation des ovocytes (Li et al, Nat Commun, 2022).
Concernant ces fibres de stress en expansion, nous explorerons trois questions principales : 1) le passage de fibres de stress contractiles à des fibres de stress en expansion ; 2) la mécanique des fibres de stress en expansion; 3) la corrélation entre les fibres de stress en expansion et l’adaptation des tissus. Comprendre les différentes fibres de stress in vivo nous permettra de mieux comprendre comment les cellules épithéliales remodèlent leurs réseaux de cytosquelette pour moduler leurs différents processus au cours du développement et de la progression du cancer.
Projet 3
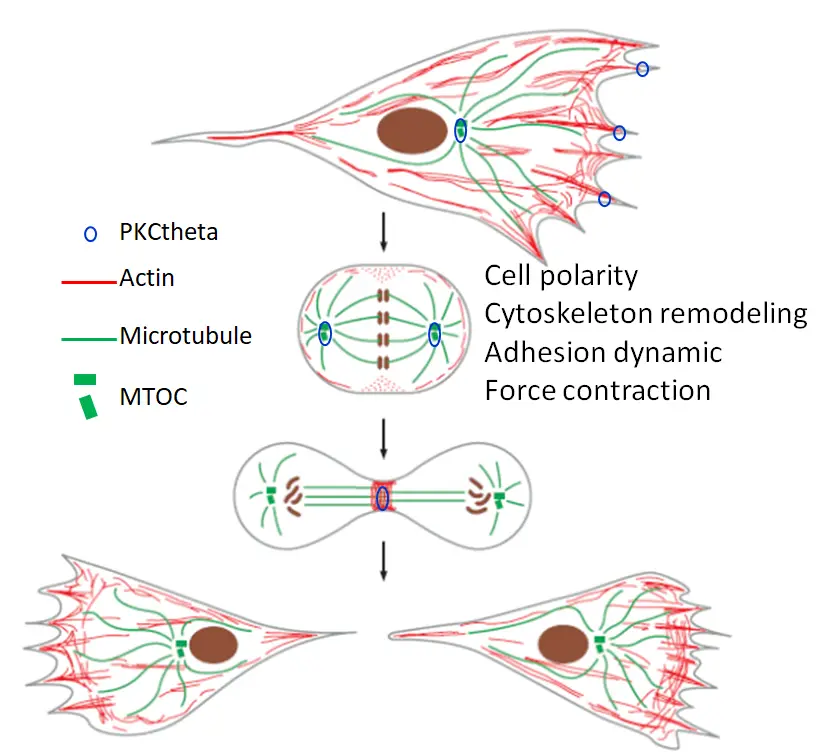
Le cancer est causé par la dérégulation des processus biologiques fondamentaux dans les cellules normales. Bien que les capacités de migration et de division cellulaires soient souvent progressivement augmentées au cours du développement et de la progression du cancer de type non invasif à hautement métastatique, le lien entre ces deux comportements cellulaires est rarement exploré. Ici, une question importante est de savoir si et comment ces deux comportements cellulaires sont intégrés par certains contrôles communs. En utilisant les cancers du sein et de l’ovaire hautement métastatiques comme modèle pour étudier l’agressivité tumorale, nous avons constaté que la sérine/thréonine kinase PKCtheta, une enzyme clé dans l’activation des lymphocytes T, est fortement exprimée dans ces sous-types de cancer particulièrement agressifs (Chadelle et al, Cancer Letters 2022). PKCtheta présente plusieurs localisations subcellulaires spécifiques dans ces cellules cancéreuses agressives impliquant diverses fonctions biologiques. En effet, PKCtheta est impliqué dans le contrôle de la prolifération, de la migration et de l’invasion des cellules cancéreuses agressives ainsi que de leur activité métastatique. Ici, notre objectif principal est de déterminer si et comment PKCtheta utilise des facteurs et des mécanismes communs tels que la polarité cellulaire, le remodelage du cytosquelette, la dynamique d’adhésion et la contraction des forces, pour gouverner et coordonner la migration et la division cellulaires. Ce comportement intégré amplifierait sans aucun doute l’impact de PKCtheta sur l’agressivité tumorale.
– Li S, Liu Z-Y, Li H, Zhou S, Liu J, Sun N, Yang K-F, Dougados V, Mangeat T, Belguise K, Feng X-Q, Liu Y, Wang X*. Basal actomyosin pulses expand epithelium coordinating cell flattening and tissue elongation. Nat Commun (2024) 15(1):3000.
– Zhou S, Li P, Liu J, Liao J, Li H, Chen L, Li Z, Guo, Q, K Belguise, Yi B, Wang X*. Two Rac1 pools integrate the direction and coordination of collective cell migration. Nat Commun (2022) 12;13(1):6014.
– Chadelle L, Liu J, Choesmel-Cadamuro V, Karginov AV, Froment C, Burlet-Schiltz O, Gandarillas S, Barreira Y, Segura C, Van Den Berghe L, Czaplicki G, Van Acker N, Dalenc F, Franchet C, Hahn KM, Wang X*, Belguise K*. PKCθ-mediated serine/threonine phosphorylations of FAK govern adhesion and protrusion dynamics within the lamellipodia of migrating breast cancer cells. Cancer Lett (2022) 526:112-130.
– Popkova A, Stone OJ, Chen L, Qin X, Liu C, Liu J, Belguise K, Montell DJ, Hahn KM, Rauzi M, Wang X*. A Cdc42-mediated supracellular network drives polarized forces and Drosophila egg chamber extension. Nat Commun (2020) 11(1):1921.
– Wang H, Qiu Z, Xu Z, Chen SJ, Luo J, Wang X*, Chen J*. aPKC is a key polarity molecule coordinating the function of three distinct cell polarities during collective migration. Development (2018) 145(9).
– Qin X, Hannezo E, Liu J, Mangeat T, Choesmel-Cadamuro V, Macdonald J, Liu Y, Yi B, Wang X*. A biochemical network controlling basal myosin oscillation. Nat Commun (2018) 9(1):1210.
– Qin X, Park BO, Liu J, Chen B, Choesmel-Cadamuro V, Belguise K, Heo WD, Wang X*. Cell-matrix adhesion and cell-cell adhesion differentially control basal myosin oscillation and Drosophila egg chamber elongation. Nat Commun (2017) 8:14708.
– Combedazou A, Choesmel-Cadamuro V, Gay G, Liu J, Dupré L, Ramel D, Wang X*. Myosin II governs collective cell migration behaviour downstream of guidance receptor signalling. J Cell Sci (2016) pii: jcs.179952.
Affiliation



