


- Institute
- Research topics
- Organization
- Platforms
- Services
- Europe/International
- Science outreach
- Agenda
- Directory
- Access

In the team, we use biophysical modeling to help us understand cell and tissue behavior during morphogenesis. This approach nicely complements our experimental work on Drosophila, chicken and quail. Indeed, while in vivo approaches could have technical limitations, biophysical modeling allow us turning on/off any components of our models at wish and dissect their respective contributions to a specific morphogenetic process.
Contribution to the creation of Tyssue: an epithelium simulation library
To model the biological processes studied in the team, we contribute to the development of an open-source Python library: Tyssue set up by Guillaume Gay from the Turing Center in Marseille. The Tyssue Python library seeks to provide a unified interface to implement bio-mechanical models of living tissues. Its main focus is on vertex based epithelium models. Tyssue allows to model the mechanical behavior of 2D, apical 3D or full 3D epithelia (Fig1) based on the numerical resolution of the equations of motion for the mesh vertices.
Link to the article: Theis et al, 2021 Joss

Modeling epithelium folding
One of our main models is the developing leg of Drosophila pupae, and more particularly the implication of apoptotic cells in fold formation.
To know more about the project: link
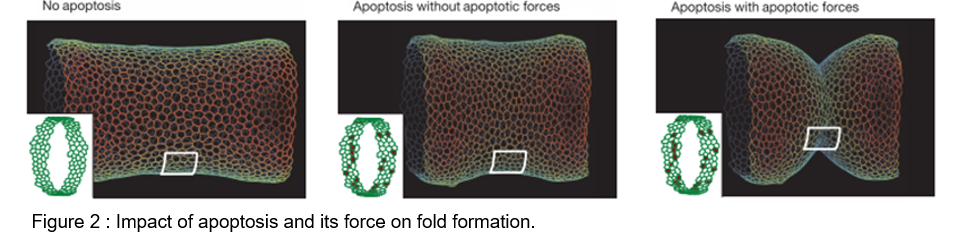
Impact of apico-basal apoptotic forces
To test the role of apoptotic forces in folding, we elaborated a physical model based on the 3D apical vertex model. In the absence of the apoptotic dependent forces, the model indicates that the simple disappearance of 30 cells from a continuous ring-like domain is not sufficient to induce cell shape reorganisation and to create an invagination at the tissue level (middle panel, compare to the left panel).
Interestingly, if a transient apico-basal pulling force is generated at the level of individual apoptotic cells and is translated into an increased contractility in the neighbouring cells, we observe a reorganization similar to the one observed in the leg fold, both at the cellular and tissue level. This shows that the added effect of both apoptotic forces is necessary and sufficient to induce folding in silico (right panel, compare to the left pannel).
Using this model, we could further test the respective roles of each of these mechanical forces: the transient apico-basal force of apoptotic cells and the apical transmission of this force to the neighbours through an increase of apical contractility. If we only apply the transient apico-basal force in each dying cell, without transmission of this force to the neighbouring tissue, an invagination response is observed all around the cylindrical tissue, albeit irregular. Now, if an increase of apical contractility is applied in two rows of apoptotic neighbours, without applying the apico-basal pulling force of the apoptotic cells, a strong reorganization of cell shapes is observed, although the tissue shape remains mainly unchanged. In conclusion, both forces are required to obtain a deep and regular fold.
Link to the article: Monier et al, 2021 Nature
Morphogenesis Robustness
We observed that developing leg folding occurs in a mechanically noisy environment. To test the impact of mechanical perturbations on fold formation, we implemented our previous model of developing leg folding and integrated random mechanical perturbations in close vicinity to the predicted fold domain, mimicking the mechanical noise observed in vivo. Interestingly, we found that mechanical noise appears sufficient to induce fold deviations in the simulations, mimicking the absence of Myosin planar polarity in the tissue (Fig3 left).
We then wondered whether restoring myosin II polarity in a non-polarized tissue could rescue the defects of fold deviation. In order to integrate myosin II planar polarity in the model, we modeled the tension associated with the myosin II planar polarity by the attribution of different values of junctional tension depending on junction orientation in our model. Fold robustness was unaffected by the integration of tension polarity in the model in the absence of external perturbations, while interestingly, gradual increase of tissue polarity favors fold straightness and insensitivity to surrounding perturbations. We further quantified fold morphogenetic robustness in our theoretical model for different degrees of polarity. Interestingly, increasing polarity decreases the degree of deviation of the fold and thus decreases the variability in fold directionality (Fig3). These data support the fact that the planar polarization of tissue tension favors mechanical isolation of fold formation, which ultimately ensures morphogenetic robustness.
Link to the article: Martin, Theis et al, 2021 Developmental Cell

Modeling mesoderm invagination
Another project in the team focuses on the impact of EMT on the surrounding tissue using both the controlled induction of EMT in epithelial tissues (imaginal discs) and the mesoderm invagination (embryo) in Drosophila.
To know more about the project : link.
EMT apico-basal forces drive epithelium remodeling
In drosophila imaginal tissues, we found that at the onset of EMT (in a controlled inducible system), cells generate myosin II-dependent apico-basal forces while constricting their apex and pull down transiently the apical surface of the epithelium, before delaminating. At the tissue scale, ectopic EMT appears sufficient to drive ectopic folding, suggesting that the force generated by predelaminating cells constitutes a driving force for tissue remodeling. We further found that similar apico-basal structure are formed in cells from the mesoderm in Drosophila embryo and that an apico-basal force appears essential for mesoderm invagination. To test specifically the contribution of apico-basal forces generated by pre-delaminating cells in tissue folding, we developed a physical model of the embryo apical junction network based on the vertex model we implemented recently. This 3D model of mesoderm invagination mimicks the cellular dynamics observed in the embryo (anisotropic constriction, gradual and asynchronous apical constriction, and apical force propagation). In this model, mesodermal cells progressively constrict their apical surface preparing their future delamination. Simulation results showed that apical tension alone without apico-basal force only leads to a curved-shaped mesoderm, independently of the strength of apical contractility. Only in the presence of apico-basal tension does the mesoderm invaginate, forming the V-shape observed in vivo (Fig 4 right panel).
Link to the article: Gracia, Theis et al, 2019 Nature Communication

Follow us 
Université Paul Sabatier
118 Route de Narbonne
31062 TOULOUSE Cedex
France
05 61 33 58 00


